第五节 睡眠与觉醒的脑机制
人的一生三分之一的时间用于睡眠,大多数人在睡眠中都有梦的体验。因此睡眠与梦的生理心理学问题,很久以前就引起人们极大关注。弗洛伊德对梦的解释曾引起人们的兴趣,巴甫洛夫的睡眠理论也广为流传。然而,人类对睡眠与梦的生理心理学机制的现代科学认识,仅仅是最近四十多年的事。20世纪60年代,借助成熟的电生理学技术找到了睡眠类型和梦的生理指标,为睡眠的研究奠定了科学基础。70年代,脑化学通路理论的成熟为睡眠与梦的研究提供了新的前景。神经解剖学、生理学和脑化学的综合研究,从器官水平、细胞水平和分子水平上加深了对睡眠与梦机制的认识。然而,关于睡眠与梦的许多问题至今仍是科学之谜,尚需深入研究。
一、睡眠类型与睡眠周期
人类的睡眠可以分为两种类型,慢波睡眠(slow wave sleep)和异相睡眠(paradoxical sleep)。在慢波睡眠中,脑电活动以慢波为主,脑电活动的变化与行为变化相平行,从入睡期至深睡期,脑电活动逐渐变慢并伴随着逐渐加深的行为变化,表现为肌张力逐渐减弱,呼吸节律和心率逐渐变慢。在异相睡眠中,脑电变化与行为变化相分离,脑电活动类似慢波睡眠的入睡期,以肌张力为代表的行为变化却比深睡期还深,肌张力完全丧失,还伴有快速眼动现象和桥脑-膝状体-枕叶PGO波周期性高幅放电等特殊变化。所以,异相睡眠又常称为快速眼动睡眠(rapid eye movement sleep,REM),这种类型的睡眠与做梦的关系比慢波睡眠更为密切。
根据脑电活动和行为变化的平行性,将慢波睡眠分为4个发展时期,睡眠一期(入睡期),行为上安静困倦开始进入睡眠状态,清醒安静状态下的脑电活动(以8~13次/秒的α 节律为主)变得不规则,α 波和不规则快波交替出现。大约10分钟以后进入慢波睡眠的二期(浅睡期),脑电活动更不规则,在4~7次/秒θ 波的背景上出现13~16次/秒的睡眠纺锤波,环境中出现意外声音,此时脑电图上可出现高幅的K复合波,代表脑电活动的短暂唤醒反应,在浅睡期中,被试已经入睡,并出现酣声,但将被试叫醒后却常自称尚未睡着。慢波睡眠二期大约持续15分钟后转入慢波睡眠三期(中睡期),脑电活动在θ 波背景上出现20%~50%的0.5~3次/秒δ 波。再经15分钟左右,慢波睡眠三期为四期(深睡期)所取代,脑电活动50%以上为高幅δ 波。处于中睡期,被试已经睡熟,但尚易叫醒,处于深睡期的被试不但睡熟还难以叫醒。慢波睡眠1~4期过渡中,颈部肌肉和四肢肌肉张力逐渐降低,心率和呼吸逐渐变慢,体温、脑温降低,闭眼、缩瞳,脑血流量较清醒安静时为多,脑下垂体分泌的生长激素和促肾上腺皮质激素以及肾上腺分泌的肾上腺皮质激素在慢波睡眠中比在白天清醒时增多。特别是生长激素,分泌的高峰在慢波睡眠的四期。在慢波睡眠各期中被唤醒后,报告做梦者人数极少。即使做梦者报告,其梦境也平淡、生动性弱,概念性和思维性较强。但梦魇或噩梦惊醒者多发生在慢波睡眠第四期。此时睡梦者醒后只能陈述恐惧感,被追捕或掉入深渊等危险境界,不能陈述梦境的全部故事情节。
在慢波睡眠之后,常出现异相睡眠。此期睡眠者肌肉呈完全松弛状态,甚至肌肉电活动完全消失,睡眠深度似乎比慢波四期更深,体温仍较低,对外部刺激的感觉功能进一步降低,难以将睡眠者从此期立即唤醒。与行为变化相反,脑电活动为极不规律的低幅快波,类似清醒期和慢波睡眠一期的脑电变化。脑的温度、脑血流量、脑耗氧量迅速增加,呼吸心率也时而突然加快,甚至一些支气管哮喘病在此期睡眠中可突然发作哮喘;心脏病人也可能发作心绞痛,内分泌活动的特点是生长激素分泌迅速降低,性腺和肾上腺皮质分泌活动增强。生殖器充血,分泌物增多或阴茎勃起、遗精等。在异相睡眠中,最有特征性的行为变化是眼球快速运动,约每分钟60次左右。正是由于这一特点,常将此期睡眠称为快速眼动睡眠。与之相应,眼电现象显着加强,在桥脑、外侧膝状体和枕叶皮层中可记录到周期性的高幅放电现象,称之为PGO波。从异相睡眠中唤醒后,80%以上的人声称正在做梦,尚可陈述梦境的故事情节,形象生动以视觉变幻为主。
人的每夜睡眠大约由慢波睡眠和异相睡眠交替变换的4~6个周期所组成,平均每个周期历时80~90分钟,包括20~30分钟异相睡眠和约60分钟的慢波睡眠。成人入睡后,必须先经过慢波睡眠一~四期和四~二期的顺序变化后,才能进入第一次异相睡眠。从上半夜到下半夜每次更替一个周期,异相睡眠的时间都有所增长。所以,后半夜睡眠中,异相睡眠时间的比例增大。整夜睡眠中各期所占时间比例平均分配是慢波二期占50%,异相睡眠占25%,慢波三、四期各占10%,慢波一期占5%。正常睡眠周期的紊乱可能导致许多特殊的病理性睡眠状态,了解和研究这些病理性睡眠状态,可以加深我们对睡眠类型和睡眠周期的认识。
发作性睡病(narcolepsy)、猝倒(cataplexy)和入眠前幻觉(hypnagogic hallucination)是异相睡眠中的常见障碍,夜尿症(nocturnal enuresis)、梦游症(somnabulism)和夜惊症(night terrors)则是慢波睡眠的常见障碍。发作性睡病又称嗜睡症,主要症状是在不应睡眠的工作时间内,突然不可控制地陷入睡眠状态,特别是在单调或枯燥的环境中更容易发作。每次发作性睡眠持续2~5分钟,醒来后觉得精神很好。可以把猝倒看作是发作性睡病的另一种表现形式。发作时全身肌肉张力突然消失,病人摔倒好像是从清醒状态突然进入异相睡眠阶段,持续几秒钟至几分钟。猝倒不同于发作性睡病,一般不会自发地发作,情绪波动是最常见的诱发因素,生气、大笑或紧张地完成某一动作,如试图抓住从身边飞过的物体等,均可引起猝倒。入眠前幻觉表现为在早上即将醒来或躺在床上刚入睡时,突然陷入异相睡眠状态,因为肌肉张力完全消失,体验到可怕的情景却呼叫不得也动不得。别人呼叫他的名字或轻轻拉动他的身体,可使之立即摆脱此种幻觉状态,恢复正常后还能描述幻觉内容与内心体验。上述三种睡眠障碍共同特点是其发作性,从清醒期越过慢波睡眠阶段突然陷入异相睡眠状态,是异相睡眠脑机制的障碍。有人发现这些睡眠障碍有家族遗传因素。苯丙胺、丙咪嗪对这类睡眠障碍有一定疗效,说明脑单胺类神经递质功能的增强对改善这类睡眠障碍是有效的。苯丙胺促进单胺类递质从突触前释放,丙咪嗪抑制突触前成分对单胺神经递质的重摄取,两者均使突触间隙中单胺类递质的浓度增高,从而增强了神经信息的传递功能。两种药物的疗效说明这类睡眠障碍与异相睡眠的脑生化机制障碍有关。与此不同,夜尿症、梦游症和夜惊症均出现于幼儿慢波睡眠四期。夜尿症病儿常在睡眠的慢波四期尿床;梦游症病儿在睡眠的慢波四期中,从床上起来进行一些刻板动作,事后又回床继续睡眠,次日不能回忆出夜间的异常行为,夜惊症病儿在睡眠的慢波四期出现惊叫、颤抖、手足快速运动等极度恐怖表现,事后对这种体验不能回忆。总之,慢波睡眠障碍与异相睡眠障碍不同,肌肉尚保持一定张力,可以进行某些动作;但事后完全不能回忆。异相睡眠障碍不伴有动作表现,且事后对梦境体验能够回忆和叙述。由此可以看出慢波睡眠和异相睡眠有不同的机制。
二、睡眠的功能与睡眠剥夺的实验研究
睡眠是生物机体的本能行为之一,与饮食行为、性行为和防御攻击行为一样,对维持种族延续和个体生存具有同等重要的意义。人们可以绝食自尽,但却无法有意识地控制自己长期拒绝睡眠。那么人为什么要睡眠,睡眠有什么功能?不言而喻,休息和从疲劳中恢复是睡眠的重要功能之一。从更积极的意义上理解,睡眠还有促进生长发育、易化学习、形成记忆等多种功能。睡眠剥夺的实验研究,对睡眠功能的认识提供了许多科学证据,加深了我们对睡眠功能的理解。
睡眠完全剥夺200小时,可能会导致人的情感不稳定、易激惹、注意力涣散、记忆减退、思维迟钝和偏执状态。迫使大鼠不停地运动,完全剥夺睡眠5~23日,会使之变得非常虚弱,运动不协调,甚至死亡。死后解剖发现,肾上腺增大、胃溃疡、肺水肿……另一方面,发现一些参加体育竞赛项目之后的运动员整夜睡眠增加18%~27%。仅仅计算慢波睡眠时间,则竞赛后明显增加至40%~45%。由此可见,睡眠对于解除疲劳、恢复体力是十分必要的。进一步分析表明,如果以脑电图出现高幅δ 波为主的活动为指标,则说明睡眠已进入慢波四期(深睡期),此时唤醒被试,使之慢波四期睡眠被选择性剥夺,结果发现四期睡眠的回跃现象(rebound phenomenon),即剥夺慢波四期睡眠之后的正常睡眠过程中,会出现更多的慢波四期睡眠。这说明在体力活动之后的恢复中,慢波睡眠四期可能更为重要。选择性剥夺异相睡眠,即每当出现快速眼动时立即唤醒被试,数日之后常使人们陷入焦虑抑制状态。在异相睡眠剥夺之后,恢复正常睡眠时也会出现异相睡眠的回跃现象,当异相睡眠得到补偿之后,被试的情绪状态也恢复正常。由此可见,异相睡眠对正常情绪状态维持具有重要意义。但是,令人不解的是抗抑郁药和电抽搐治疗虽对治疗抑郁症十分有效,却同时抑制了异相睡眠,这与上述设想相矛盾。因此,近些年异相睡眠与情感活动间的关系又引起人们的怀疑。中国科学院心理所李德明利用小站台水环境技术,对大鼠选择性剥夺异相睡眠,让大白鼠立于小站台上,一旦它们陷入异相睡眠,由于肌张力完全丧失而落入四周的水中,从而又惊醒动物。首先对大鼠进行灯光回避辨别反应模式的训练,然后以不同时间间隔,应用小站台水环境技术剥夺异相睡眠。结果表明,大鼠在学习后立即剥夺异相睡眠24小时对记忆巩固明显不利;如果在立即剥夺异相睡眠24小时之后,再让其恢复自由睡眠24小时,记忆成绩无明显改善;反之,学习后先让动物自由睡眠24小时,再行剥夺异相睡眠24小时,则记忆不受影响。这一结果证明,异相睡眠对于学习后一段时间内记忆的巩固具有重要意义。他们进一步研究发现,虽然异相睡眠剥夺对回避反应的短时记忆和记忆保持有明显的影响,但对动物回避反应的学习能力则没有影响。国外研究也表明异相睡眠对记忆形成的影响不是“全或无”的,异相睡眠剥夺只能延迟或影响记忆的形成,但不能完全妨碍记忆的形成。此外,给动物较多的学习任务,则随后就会出现较多的异相睡眠,一旦习得行为巩固,则异相睡眠的时间比例也会恢复常态。异相睡眠与记忆形成的这种关系,可能与此时脑蛋白质合成率较高有关。
睡眠过程中脑下垂体分泌的生长激素增高,在整夜睡眠的第一个慢波睡眠4期出现时达高峰,随后生长激素沿血液循环达全身各处发挥生理作用。这恰好处于慢波睡眠4期之后的异相睡眠期。躯体组织各种细胞,特别是儿童骨骼细胞迅速分裂,蛋白质合成率也相应地迅速增加。这说明睡眠有助于未成年机体的生长发育,此外,还有人认为异相睡眠中,蛋白质合成率增加可能与睡眠之前受到各种刺激的信息编码和记忆储存有关。对整夜睡眠的梦分析表明,每夜睡眠中第一、二两次异相睡眠的梦多以重现白天的活动内容为主,似乎对当天经历进行着重新整理和编码;第三、四两次异相睡眠的梦多重现过去的经历甚至是儿时的体验;第五次异相睡眠的梦既有近事记忆又有往事记忆的内容。这些事实似乎支持异相睡眠中蛋白合成增加与信息编码、短时记忆以及长时记忆储存有关。有人认为儿童期接触很多新事物,需要编码和储存的信息较多,睡眠较多;老年人需要学习内容减少,睡眠也减少。然而,实验研究发现,在4岁以下的幼儿和许多老年人的睡眠中,在慢波四期和异相睡眠期,生长激素的分泌与蛋白质合成率并未增多。所以,异相睡眠与学习记忆和脑代谢的关系尚需进一步深入研究。
比较心理学家们考查了各种动物的睡眠规律,认为睡眠是一种生态适应性反应,使有机体在漫长黑夜无法觅食时能安静地睡在一处以减少体力消耗和避免敌人发现。这种假说不能很好解释人类睡眠的意义和功能。从灵长类进化为人类经历了几十万年,人类能制造食物和建筑房屋,但睡眠规律并未发生重要变化。所以,这种假说未必可靠。
三、经典睡眠理论
关于睡眠与觉醒机制,巴甫洛夫在20世纪初,就较早地从神经生理学角度提出主动性内抑制的睡眠理论。20世纪50年代通过着名的孤立头与孤立大脑标本和电生理技术,搞清了在维持觉醒中脑干网状结构的重要作用;60年代弄清了睡眠的类型和睡眠周期,这些构成睡眠的经典理论。
(一)巴甫洛夫的睡眠理论
八十多年前,巴甫洛夫在研究动物条件反射的实验室中,发现对狗进行分化抑制的条件反射训练,可使狗陷入睡眠状态。在大量实验资料的基础上,他提出了一套睡眠理论,其基本要点在于睡眠的本质是大脑皮层起源的广泛扩散的内抑制;这种抑制在皮层中和向皮层下脑结构扩散过程中存在一定的时相,构成从觉醒到完全睡眠的过渡,即催眠相;梦是由于内外环境因素的影响,在普遍抑制背景上大脑皮层细胞群局部的兴奋活动。
睡眠和内抑制是同一过程,消退抑制、延缓抑制、条件抑制和分化抑制等诸种内抑制不仅与睡眠可以相互转化和替代,这些抑制的总和也可以导致睡眠。睡眠和内抑制有同样的发展和运动规律,即扩散与集中和相互诱导的规律。小孩在入睡前常出现兴奋增强期,表现为哭闹不止,即为睡眠抑制正诱导现象。睡眠和内抑制也有许多不同之处。内抑制是在觉醒状态下,个别皮层细胞群的抑制,是分散的、局部的抑制过程;睡眠则是广大皮层区、皮层下脑结构直至中脑的广泛性抑制过程。睡眠时不能保持直立的姿势,肌肉张力大大降低,说明抑制过程波及到中脑以下运动系统的功能。睡眠抑制在脑内并不均匀,常常存在某些易兴奋点在警卫着睡眠,哺乳期的母亲在熟睡中不能为雷鸣般巨大声响所唤醒,却极易为婴儿的啼哭声所唤醒。睡眠中脑抑制的不均匀性还成为梦的基础。在广泛抑制的背景上,某些脑细胞群摆脱抑制而兴奋起来,产生了梦的现象。梦的内容可以反映出内外环境刺激因素的性质,睡眠时身体不舒适或心肺功能不畅,常出现噩梦;膀胱充盈常有到处寻找厕所的梦境等。
巴甫洛夫的睡眠理论不仅阐明了睡眠的本质、睡眠的起源和一些常见的睡眠现象如梦等的生理基础,还揭露了从清醒到深睡之间催眠相,并以此为根据,解释了多种神经精神病的病理机制,还提出了睡眠的保护性医疗作用。
在正常清醒状态下,条件反射的强弱与刺激强度间存在着一定的关系;但从清醒到睡眠的过渡期内,这种强度关系发生了变化。根据强度关系的不同,巴甫洛夫将催眠相分为正常相、均等相、反常相、超反常相和抑制相等5个时相。强刺激引出强反应,弱刺激引出弱反应,阴性刺激不引起反应,这是正常相的强度规律;强刺激和弱刺激引出同样的反应,阴性刺激不引起反应是均等相的强度规律;在反常相中,强刺激引出弱反应,弱刺激引出强反应,阴性刺激不引起反应;在超反常相中,无论是强刺激还是弱刺激均不引起反应,相反,阴性刺激却引起反应;最后,在抑制相中各种刺激均不引起机体的反应,机体进入完全睡眠状态。正常人的睡眠过程,从正常相到抑制相的各催眠相发展是很快的,有些催眠相很难观察到,特别是从睡眠到清醒的过渡时,催眠相更难以观察。但是,在许多病理条件下,大脑停滞在某一催眠相可达数月或数年之久。例如,精神分裂症紧张状态的病人存在着违拗症状,让其伸手而缩回,反之,让其缩回手则伸出,类似于超反常相。
巴甫洛夫的睡眠理论形成于20世纪10~20年代,虽然他天才地概括出睡眠发展过程中脑的宏观生理机制,并解释了许多与睡眠有关的日常性和病理性现象。然而,由于历史的局限性,他不可能从神经细胞水平上和分子水平上进一步揭露睡眠的本质。60年代确立的现代神经生理学,用电生理技术揭露了在睡眠起源于皮层下脑干网状结构的重要发现,使人类对睡眠与觉醒机制的认识前进了一大步。
(二)脑干网状结构在睡眠与觉醒中的重要作用
1937年着名生理学家布雷默(Bremer)建立了猫的孤立脑(isolated brain)标本和孤立头(isolated head)标本。前者在中脑四叠体的上丘和下丘之间横断猫脑,此后猫陷入永久睡眠状态;后者在脊髓和延脑之间横断猫脑,则猫保持正常的睡眠与觉醒周期。他以此证明在延脑至中脑的脑干中,存在着调节睡眠与觉醒的脑中枢。1949年意大利着名电生理学家莫鲁齐(Moruzzi)和马古恩(Magoun)发现,电刺激脑干网状结构引起动物的觉醒反应。此后大量实验研究表明,无论是各种外部刺激还是感觉通路的电刺激,均沿传入通路的侧支引起脑干网状结构的兴奋,然后再引起大脑皮层广泛区域的觉醒反应。因此,把脑干上部的网状结构称为上行网状激活系统(ascending reticular activating system)。微电极电生理学技术的应用,也积累了许多科学事实,证明脑干上行网状激活系统的神经元单位活动可受多种刺激的影响,提高其发放频率。行为的觉醒水平、脑电图觉醒反应与脑干网状上行激活系统的神经元单位发放频率之间存在着确定的一致关系。这些事实证明脑干网状结构在睡眠与觉醒的机制中具有重要作用。这种认识从20世纪50年代起成为一直公认的理论;然而,80年代初的一些实验研究却提出了一些相反的事实,表明在自由活动状态下,脑干网状结构的细胞单位活动与唤醒水平或觉醒与睡眠周期之间并无直接关系。脑干网状结构的许多神经元分别与眼、耳、脸、头、躯体和四肢的局部运动有关。在过去的实验研究中,由于动物处于固定或约束的体位,又有麻醉剂的作用,个别单位活动与运动间的特异关系无法揭露出来。此外,传统研究多以细胞外电极记录出许多神经元单位活动的总和,从而使个别神经元反应特异性受到掩盖。虽然有这些相反的研究报道,迄今仍将脑干上部的网状上行激活系统看成是维持觉醒的重要脑结构。
关于脑干网状结构在睡眠与觉醒中重要作用的研究,20世纪50年代末又取得了新的进展。在桥脑中部横断脑后,发现猫绝大部分时间内(每日的70%~90%时间)处于觉醒状态,这说明桥脑中部以下的脑干网状结构对睡眠具有重要作用。麦格尼(Magni)等人将脑干上部和下部的脑血供应分离开,给脑干上部注入麻醉剂引起动物陷入睡眠状态;但将麻醉药注入脑干下部,则使睡眠的动物很快觉醒。这说明脑干下部正常对睡眠是重要的。当用麻醉剂使其活动减弱,动物的睡眠就会中止;相反将脑干上部的上行激活系统麻醉之后,由于脑干下部功能亢进,动物就会睡眠。正常状态下,脑干上部和下部之间对睡眠与觉醒的变化有相反的作用。现将20世纪60年代以前关于脑干网状结构在睡眠与觉醒中作用的研究结果以图8-2加以总结。脑干以上横断脑(孤立脑标本),动物陷入永久睡眠状态,脑干中间横断脑(桥脑中部横断),动物70%~90%时间处于觉醒状态;脑干下位横断脑(孤立头标本),动物维持正常的睡眠与觉醒周期。脑干上部的网状上行激活系统对维持觉醒状态起重要作用;桥脑下部的网状结构对睡眠起重要作用;脑干上部与下部的网状结构相互作用维持正常的睡眠与觉醒周期。这就是20世纪60年代对睡眠机制的认识水平。
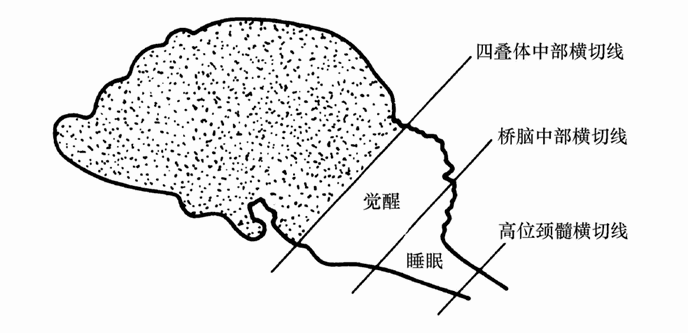
图8-2 睡眠觉醒中网状结构作用的示意图
四、对睡眠机制的现代认识
20世纪70年代以来对睡眠机制的研究已经积累了相当多的科学事实,证明脑内存在着一些关键性结构,其生理、生化过程的维持与转换对睡眠具有重要作用。对于慢波睡眠来说,关键性脑结构是缝际核、孤束核和视前区、前脑基底部;对于异相睡眠,关键性脑结构是桥脑大细胞区、蓝斑中小细胞、外侧膝状体神经元和延脑网状大细胞核等许多脑结构。与睡眠有关的化学物质是单胺类神经递质、胆碱类神经递质和多肽,特别是诱导睡眠肽(DSIP)和γ -氨基丁酸受体蛋白质。
将猫脑缝际核80%~90%神经元损毁,则使之数日内处于不眠状态,几日后虽有睡眠,但睡眠时间非常短。用对氯苯丙氨酸(p -arachlorophenylalanine,PCPA)抑制脑内5-羟色胺(5-HT)的合成过程,动物也不再睡眠。由此可见,无论是破坏缝际核还是抑制缝际核5-羟色胺神经元合成5-羟色胺的生化过程,均导致不眠状态,证明缝际核的5-羟色胺神经元在慢波睡眠中具有重要作用。
孤束核位于延脑,是味觉和内脏感觉神经核,低频电刺激孤束核,引起猫脑电活动的同步化,出现低频高幅波,并伴有睡眠的行为表现。刺激内脏或迷走神经也引起脑电活动的同步化。饱食之后,胃内过度充食易导致睡眠,这可能就是由于胃刺激沿迷走神经传入,引起孤束核兴奋的结果。
视前区位于下丘脑的视交叉之前的部位,视交叉上核是生物钟的“起搏点”,对慢波睡眠至关重要。损毁视前区使动物失眠,数日后陷入昏迷至死亡。大脑半球基底部的前脑区在慢波睡眠中也有重要作用,电刺激此区30秒后,引起大脑电活动的同步化,随后出现睡眠行为。
5-羟色胺是一种单胺类神经递质,虽然在睡眠机制中具有重要作用,但却不是与睡眠相关的特异性物质。许多生理学家在研究慢波睡眠的关键性脑结构的同时,致力于寻找脑内的特异性睡眠物质。哈佛大学医学院的蓓本海姆(J.R.Pappenheimer)等人报道,从睡眠剥夺的山羊脑脊液中提取一种睡眠因子(sleep factor),将其注入大白鼠脑室内,导致大白鼠活动减少,睡眠增加。20世纪70年代末,一些实验室分别从睡眠48小时以上的山羊、绵羊、兔和小牛等动物的脑脊液中提取出睡眠因子,将之分别注入大白鼠、猫、兔的脑室里,发现它们的睡眠都增加。睡眠因子注入后,动物慢波睡眠期增加,脑电图出现较多的高幅δ 波。经化学分析表明,它是一种相对分子质量为350~500的多肽,其作用持续时间可达10小时,发生最大作用的时间是注入脑室后的2小时。这个实验室从4.5吨重的人尿中提取30毫克睡眠因子,经化学分析表明它是二氨基酸,通常是人肠道内细菌代谢产物。因此,睡眠因子是否是脑内产生的特异性睡眠物质的问题又重新提了出来。
蒙尼尔等人(1977)在脑化学、电生理学和动物睡眠行为的综合研究过程中,提纯和确定了一种内源性睡眠肽的化学物质及其生理作用,并将之定名为“δ -诱导睡眠肽”(delta sleep inducing peptide,DSIP)。他们用低频电刺激兔的丘脑中线核,使之大脑皮层电活动出现同步化的纺锤波,此时兔脑即会合成较多的DSIP进入血液中。将该兔血液注入另一只兔脑内,就会使后者脑电活动出现同步化纺锤波。他们的研究指出,DSIP是一种9肽,只有肽链第5位天冬氨酸的氨基在α 位时,DSIP才具有诱导睡眠效应的功能。我国生理心理学家刘世熠发现DSIP肽链第5位的天冬氨酸为苯丙氨酸所取代,也具有诱导睡眠作用。
弱安定剂如安定等具有轻微的镇静安眠作用。对这类药物作用机制的研究发现,它们是通过γ -氨基丁酸GABA类神经递质受体而发生作用的。将GABA受体用药物阻断后,安定等药物就失去了镇静安眠作用。GABA受体是大分子蛋白质,因此,除了单胺类、肽类物质外,大分子的蛋白质可能也与慢波睡眠有关。
异相睡眠的脑机制比慢波睡眠更复杂,这是由于它包含的生理心理成分较多,如眼动、PGO波、肌张力完全丧失,心率呼吸改变和生动的梦境体验等。一般说,脑高位的一些关键性结构与脑电去同步化快波的呈现、PGO波发放和眼动有关;脑干低位的一些下部关键性结构与异相睡眠中的肌张力变化有关。
桥脑大细胞区散于桥脑网状结构之中,最大的神经细胞体直径可达75微米,非异相睡眠时没有单位发放,一旦动物进入异相睡眠状态,桥脑大细胞开始活动并逐渐增加单位发放频率,最高发放每次可达200~300个神经冲动。每一串单位发放都伴随眼动和PGO波发放。此时大脑电活动去同步化,出现低幅快波,肌肉张力完全消失。因此,把桥脑大细胞视为异相睡眠的开关细胞(the cellular on-switch for dreaming sleep)。与此相反,在脑干背部的蓝斑(locus coerleus)内存在许多很小的去甲肾上腺素能神经元,产生低频的单一频率发放,在慢波睡眠时,它们的单位发放频率逐渐变慢,一旦进入异相睡眠,它们的单位发放立即停止或迅速降低。因此,将蓝斑中这种小细胞称为异相睡眠的“闭细胞”(off cells)。这种闭细胞以去甲肾上腺素作为神经递质,当动物进入睡眠时,蓝斑闭细胞的去甲肾上腺素含量逐渐下降,在异相睡眠阶段含量最低;但是将动物从异相睡眠中惊醒时,则蓝斑小细胞的去甲肾上腺素却突然增高。除了蓝斑内的闭细胞,在蓝斑核内和它的四周还存在一种乙酰胆碱能神经元,异相睡眠时,其细胞单位发放率增加,由它们发出轴突达延脑网状结构的下行抑制细胞,引起其活动,从而产生下行性抑制效应,使异相睡眠时肌肉张力完全消失。延脑网状大细胞核(nucleus reticularis magnocellularis in the medulla)在异相睡眠时变得更活跃并与PGO波和快速眼动现象同时发生。这种细胞的轴突达脊髓运动神经元,与之形成抑制性突触。所以,在异相睡眠时,这种细胞的兴奋引起肌肉张力消失。记录外侧膝状体在异相睡眠中的PGO波时发现,与眼动方向同侧的膝状体PGO波大于对侧膝状体的波幅。如果两眼向右运动时,右侧的膝状体内PGO波大于左侧膝状体的PGO波。进一步分析发现,膝状体的PGO波的差异尚在眼球运动之前即可表现出来。因此,记录左右外侧膝状体内PGO波的差异,可以很快预测异相睡眠时眼动的方向。据此认为,外侧膝状体具有异相睡眠眼动的命令功能,实现着眼动方向读出的神经信息编码功能。综上所述,间脑水平的膝状体和桥脑网状大细胞与异相睡眠的启动、眼动方向和PGO波发放有关;蓝斑小细胞与异相睡眠停止有关;蓝斑内及其周围的乙酰胆碱能神经元和延脑网状大细胞核与异相睡眠时肌张力的消失有关。在与异相睡眠有关的结构中,发挥作用的化学物质有去甲肾上腺素、乙酰胆碱,还可能有γ -氨基丁酸。与慢波睡眠有关的化学物质是5-羟色胺、睡眠肽和γ -氨基丁酸的受体蛋白质。尽管当代神经科学对睡眠机制的认识已经取得了这些成果,但动物和人为什么会有睡眠和睡眠的周期性变化,至今仍未能得到准确的答案。生物钟本身就是悬而未决的巨大课题。
五、睡眠周期与生物节律
人类夜间睡眠中每个周期大约为90~100分钟,猫的睡眠周期约为25~40分钟,大白鼠的睡眠周期为6~10分钟。这种关系说明随脑的大小和神经元多少,睡眠周期按比例地发生变化,可能与神经系统活动的生物节律有关。一种假设认为异相睡眠的开细胞和闭细胞之间某些特殊物质的传输时间与睡眠周期有关。猫脑内异相睡眠开细胞和闭细胞间距离约25毫米,计算两者之间蛋白质传输时约为36分钟,恰与猫睡眠周期相似。但是,这种设想还需要更多的实验证据。
最近30年对于睡眠与觉醒周期的生物钟的研究已取得较大的成果,认识到下丘脑的视交叉上核(suprachiasmatic nucleus)起着重要的作用。大白鼠的视交叉上核由约1万个直径5~15微米的小神经元构成,总体积为0.05立方毫米,它接受视网膜发出的部分传入纤维,也接受从大脑视皮质来的传出纤维,光暗信息由这些纤维传至视交叉上核以及邻近的下丘脑视前区。该核的传出纤维主要是传到下丘脑-垂体结构,以调节神经内分泌的周期变化;该核的传出纤维也止于脑干和脊髓,以调节多种生理功能的周期性变化,它调节多种内分泌功能的周期变化,如性激素、肾上腺皮质激素、生长激素、促甲状腺激素等,还调节体温的周期变化、饮食行为周期性变化和觉醒与睡眠周期的变化。破坏双侧视交叉上核后的几周内,大白鼠24小时睡眠总量不变;但觉醒与睡眠周期却发生了明显的变化。也有人发现视交叉上核的损毁对睡眠周期的影响,主要表现为慢波睡眠周期破坏,而不影响异相睡眠。异相睡眠周期可能主要决定于桥脑网状大细胞核内的开细胞和蓝斑闭细胞的周期性变化。鲁萨克(B.Rusak)等人发现,电刺激视交叉上核对睡眠的影响,决定于刺激的时间。如果电刺激出现在动物活动期间的前一阶段,则延长动物觉醒的活动期,使睡眠推迟出现;相反,电刺激发生于动物活动期的后一阶段,则缩短活动期,使动物较早进入睡眠状态。
睡眠周期无疑是生物节律的组成部分。正如人类对生物节律的认识一样,对睡眠周期的认识仍不很完美。上面综述的科学事实表明,视交叉上核作为多种生物节律调节的重要中枢,自然地影响着睡眠与觉醒周期,主要作用于觉醒和慢波睡眠间的节律关系;对于两种类型睡眠间的周期变化更多地决定于异相睡眠“开细胞”与“闭细胞”间机能关系的周期性变化。应该说这种认识还需要更多科学证据加以支持。








本书评论