第二节 摄食行为
人和动物之所以摄食,是因为饥饿感在驱动着机体。饥、饱之感产生的生理机制是本节的中心论题。概括地讲,血糖含量的下降是引起饥饿感的原发性因素,它作用于脑和肝脏中的葡萄糖感受器,激发了脑内饥饿中枢的下丘脑外侧区兴奋产生主观的饥饿感;当机体摄食以后,消化道得到充盈,消化道内的机械感受器和葡萄糖感受器受到刺激。在食物消化吸收过程中,引起血糖升高,使脑和肝内的葡萄糖感受器,特别是脑内饱中枢下丘脑旁室核和围穹窿区的兴奋,引起饱感停止摄食行为。在饥饿感产生的机制中,包括中枢环节、化学环节和外周器官的参与。现将这些环节的相互关系总结成如下示意图:
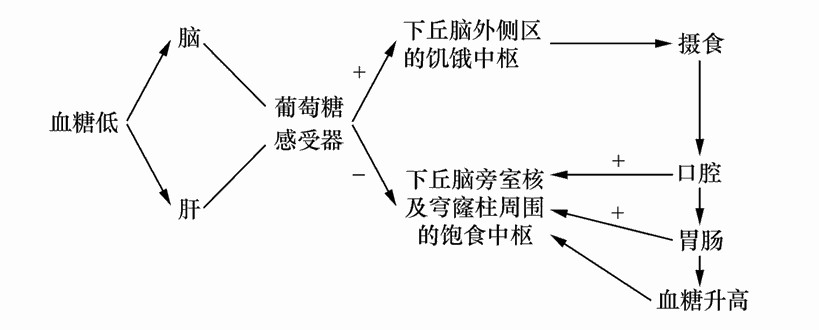
饥饱感与摄食行为的调节机制示意图
为什么这些脑结构是饥、饱的生理机制的重要中枢呢?一方面,由于这些脑结构与脑内化学通路有着交错的关系;另一方面,它们与复杂的体液调节机制也有复杂关系,与多种激素和葡萄糖代谢有关。
一、脑内的化学通路与摄食行为
下丘脑汇集了脑内的多种化学通路,现在已知的有多巴胺黑质-纹状体通路,多巴胺中脑-边缘通路,多巴胺正中隆起-垂体通路,背侧去甲肾上腺素通路,腹侧去甲肾上腺通路,5-羟色胺通路(与去甲肾上腺素通路相平行),下丘脑-垂体P物质神经末梢,下丘脑-垂体内啡肽神经末梢以及视前区、乳头体等下丘脑的乙酰胆碱神经末梢。这些化学通路对下丘脑在饥、饱调节功能中发生不同的作用。
多巴胺黑质-纹状体通路穿越下丘脑外侧区,利用电损毁或6-羟多巴胺选择性损毁这些纤维,均可使动物陷入不饮、不食状态。利用α -对甲基酪氨酸(α -methyl-p -tyrosine)抑制多巴胺的生物合成过程,也会引起同样的效应。相反,多巴胺受体激动剂则可使动物恢复饮食行为。微电极电生理研究发现,饥饿时动物脑内黑质的多巴胺神经元单位发放频率加快;对动物静脉注入葡萄糖可以降低这些神经元的单位发放频率。这一结果证明多巴胺神经元活动与饥、饱感有密切关系。
下丘脑旁室核含有丰富的去甲肾上腺素能神经末梢。动物摄食时,此核的去甲肾上腺素含量最高。由于去甲肾上腺素在这里发挥抑制性神经递质作用,所以对旁室核进行去甲肾上腺素灌流,则发现其神经元单位发放频率降低,同时诱导出动物的过食行为。由去甲肾上腺素诱发的这种过食行为在切断支配胰腺的迷走神经之后立即消失,说明旁室核中去甲肾上腺素神经系统引起的过食行为与其对胰岛分泌功能的影响有关。去甲肾上腺素之所以具有增强进食行为的效应,还可能与内侧前脑束对动物阳性自我刺激行为的强化作用有关,因为内侧前脑束也以去甲肾上腺素为神经递质,它在从中脑向前脑的上行途中穿越下丘脑外侧区。所以将去甲肾上腺素注入下丘脑外侧区也能增强食欲,使动物摄食行为增强。进一步研究发现,去甲肾上腺素对动物进食的增强效应是通过它们的受体实现的。旁室核内α 受体分布密度较高,下丘脑围穹窿区β 受体分布较多。去甲肾上腺素的α 受体激活引起饥饿感,β 受体激活引起饱感。去甲肾上腺素与α 受体结合引起进食行为,α 受体阻断剂则能终止进食行为;去甲肾上腺素与β 受体结合引起饱感并停止进食,β 受体阻断剂则增加进食。所以,去甲肾上腺素在脑的作用部位不同,对摄食行为的作用效果不一。当去甲肾上腺素在旁室核内减少,而在下丘脑穹窿区增多时就会引起饱感并停止进食。
5-羟色胺通路在脑内与去甲肾上腺素通路的分布相平行,两者的生理效应一般是拮抗的。去甲肾上腺素引起进食增强效应,而5-羟色胺则对摄食行为有抑制效应。一种引起厌食的药物芬氟拉明(fenfluramine)正是通过其增强5-羟色胺的释放而发生作用的。如果将脑内缝际核的5-羟色胺神经元事先损坏,则芬氟拉明的厌食效应就不会发生。对氯苯丙氨酸(PCPA)抑制色氨酸羟化酶活性,使脑内5-羟色胺合成减少,将其注入大鼠下丘脑,则引起动物饮食过量和体重明显增加。
除上述3种主要单胺能神经通路外,下丘脑的P物质能神经末梢、内啡肽能神经末梢和乙酰胆碱能神经末梢都参与摄食行为的调节,与相应受体结合引起摄食增多的效应。这几种脑内活性物质的受体拮抗剂均可阻断它们的摄食效应。
二、体液调节机制
除了脑内的化学通路,在脑和消化道内还存在着许多体液机制,对中枢和外周器官发生调节作用。葡萄糖及其感受器、胰岛素、胰高血糖素、肾上腺皮质激素、胆囊收缩素和垂体分泌的激素,在摄食行为的调节中均有一定的作用。与脑内化学通路的作用方式不同,这些物质随血液运行,通过脑血流作用于与摄食行为有关的脑结构而发挥生理效应。与脑内化学通路相比,这些物质作用的距离远,发挥生理效应的环节多,所需的时间较长。
肝和脑内均存在着葡萄糖感受器,对血液内葡萄糖含量进行灵敏的检测。下丘脑外侧区的葡萄糖感受器对低血糖敏感,引起饥饿感;旁室核和下丘脑围穹窿区葡萄糖感受器对高血糖敏感,产生饱感。旁室核的神经冲动沿轴突传至脑干迷走神经运动背核,产生抑制效应。迷走神经对胰腺的兴奋作用,使胰岛细胞分泌较少的胰岛素。旁室核的兴奋还合成较多的神经激素——促肾上腺皮质激素释放因子(CRF),CRF沿垂体门脉系统的血液循环作用于垂体前叶,促使其释放促肾上腺皮质激素(ACTH)。血液中的ACTH作用于肾上腺皮质,促使其释放肾上腺皮质激素(考的松)。由考的松通过肾上腺髓质再作用于胰腺,使胰岛细胞减少胰高血糖素(glucagon)的分泌。这样,在胰岛细胞中分泌的两种激素——胰岛素和胰高血糖素相互制约,调节着血糖的浓度,而两种激素的分泌又由下丘脑摄食中枢通过神经-体液机制加以调节和控制。如果血糖低,胰岛素含量高,则动物就会出现过度摄食行为;反之血糖高,胰高血糖素也高,则动物就会出现厌食行为。下丘脑腹内侧核的损毁,将旁室核与迷走神经运动背核的联系切断,使迷走神经运动背核失去抑制而过度兴奋,分泌较多胰岛素。与此同时,旁室核的兴奋却可以正常地按神经激素的许多环节作用于胰岛,使之减少胰高血糖素的分泌。由于胰岛素含量高,经消化道吸收的葡萄糖就会立即转化为贮存的形式——肝糖原、肌糖原和脂肪。高胰岛素也妨碍血液葡萄糖作为能源加以利用。因而,下丘脑腹内侧核损伤,就导致过食和肥胖的后果。
体液调节中的另一个重要物质是由十二指肠分泌的胆囊收缩素(CCK),它既作用于消化道,又可随血流作用于脑,故又称为脑-肠肽,更确切地说胆囊收缩素是脑肠肽的一种,每当十二指肠从胃内接受食物时,就会分泌CCK。血液内CCK的浓度与十二指肠从胃内接受的营养多少有关。如果营养充足,血内浓度较高的CCK就抑制胃的排空,同时随血液作用于脑内“饱中枢”,引起饱感,使机体停止进食。只有当机体吃了一定食物,并当食物从胃内大量进入十二指肠时,CCK才对进食行为有抑制作用。所以,它总是快吃饱时才发生对进食的抑制作用,饥饿者刚刚进食时,CCK就没有这种作用,说明CCK是具有饱感信号性质的物质。实验证明,切断迷走神经胃支,CCK的这种作用消失;但切断迷走神经肝脏支和胰腺支则不妨碍CCK的这种作用。这说明CCK的作用除有体液调节机制外,还有神经传导的途径。CCK作用于胃平滑肌改变其收缩程度的同时引起平滑肌中感受器兴奋性的变化,向脑内输入产生饱感的神经冲动。由此可见,一些体液调节机制,与外周消化道在饥、饱感中的作用有着密切关系。
三、外周作用与习惯
从CCK的作用机制中,可以看到胃、十二指肠在饥、饱调节中的作用。一些生理心理学家采用消化道手术方法分别考查了消化道不同部位对饥、饱感的影响。在胃幽门处手术植入一线套,对比拉紧线套前后动物摄食行为的变化,即比较阻断胃和十二指肠前后动物的摄食行为。结果发现在阻塞通道之前,动物有正常的摄食行为,适量食物产生饱感后,动物停止进食。如果拉紧线套使食物停滞于胃内,再从胃内抽出10毫升食物,则发现动物又会吃进8毫升食物。此时切断动物迷走神经胃支,使胃内的神经冲动无法传向大脑,从胃内提出10毫升食物,则动物就会过量进食使胃受到过分扩张,这些结果说明,只要十二指肠以上的口腔和胃受到食物刺激就会产生饱感,不需要小肠以下消化道的参与,但此时胃的神经支配必须保持正常。在饱感形成中,胃的作用可能与其对味觉的影响有关。胃充盈往往使味觉神经元对食物味道的反应不灵敏;反之,饥饿时胃排空,味觉神经元对食物的反应就比较强。所以,胃充盈时产生饱感的同时,对食物味道反应也变差。为了证明胃扩张在饱感和食物味感中的作用,生理心理学家们还在胃内植入气囊,用气体扩张胃,重现了食物充盈胃的效果。除了胃之外,口腔也是重要的,许多食道癌或食道狭窄的病人进行胃瘘手术,将食物直接注入胃中。这些病人的共同体验是必须将食物放入口腔咀嚼,然后将之吐入胃瘘管,注入胃中才能产生理想的饱感。对动物实行食道切断手术,使之从口腔吃进的食物不能进入胃内而落入外面。这种假饲实验证明,仅仅经过口腔咀嚼,动物也会产生短暂的饱感:但终因胃内空空,很快又去进食。这些资料证明口腔咀嚼和胃的充盈在饱感的产生中具有重要作用。
十二指肠对饱感的影响主要是通过CCK体液调节机制而实现的。此外,十二指肠将食物中的营养吸收到血液中,再经肝门脉系统转移至肝内。肝脏中的葡萄糖感受器对饱感的产生也具有一定意义。应用不能透过血脑屏障的糖,例如,果糖等注入肝门脉,虽不能直接作用于脑中枢,但动物也会产生饱感,停止进食行为,但是切断肝脏的迷走神经,再向肝门脉灌流果糖,则动物就不会出现饱感。这说明肝脏在饱感形成中的作用不仅是体液性的,也包括向脑中枢传导神经冲动的神经机制。
在人和高等动物的摄食行为中,生活习惯、学习机制往往也有重要作用。每日规律地定时摄食,就会形成饥、饱感的周期变化。因肥胖不得不节制饮食的人也有共同的体验,在节食之初,因每餐不能吃得很饱常感到不舒适;以后就会习惯于胃不全扩张状态,于是饱感的标准也发生了变化。经验、习惯和学习机制对人类饮食行为的影响,是由脑高级部位的参与而实现的。颞叶皮层、边缘系统的扣带回和杏仁损伤,常常引起过食行为,前额叶皮层损伤以后,新的饮食习惯无法建立。大白鼠大脑边缘系统和杏仁核损伤以后,习得性摄食行为破坏,不能区别出熟悉的食物和不熟悉的食物,甚至有毒的食物也不加选择地食入。因此,习得性摄食行为与本能性摄食行为相比,有更高级的、更复杂的脑机制。








本书评论