5 压力和创伤之下的大脑
海军下士“胡里奥”正在为他的第一次战斗出征做准备。当时,我在为他所在的排做MMFT培训。我很快就被他的责任心和好奇心打动了。
胡里奥所在部队的一些军官对MMFT训练表现出了抵触情绪,这种态度也影响了胡里奥所在排的一些人。在这种情况下,我给40名海军士兵做了培训,他们不是在军事训练中摆出愚蠢的姿势,就是和他们的伙伴比赛打嗝或放屁;其他人拒绝和我有眼神交流;还有一些人则以过于礼貌的态度对待我,然后在他们觉得我听不见的时候拿MMFT训练开玩笑。换句话说,在最初的几周里,当这群不成熟的家伙被要求做一些他们不愿意做的事情的时候,他们常常会表现出神气活现、闷闷不乐、敷衍和炫耀自己优势的行为。
相比之下,身为排长的胡里奥则尽心尽职地让他的战友们对我给予尊重,认真听讲。他专心聆听,提出了一些非常好的问题,并且很快就将授课内容与海军工作的方方面面联系了起来。胡里奥长期习武,因此他也能轻松完成MMFT的练习。每节课后,他都会私下找我讨论所学内容的作用。
在第四阶段,MMFT参与者学习了僵硬反应,以及它在人类防御反应层级中的位置。我播放了动物和人类遭遇僵硬反应的视频片段,以便参与者观察僵硬反应是如何出现的。在讨论过程中,参与者们也常常会讨论他们之前经历过的僵硬反应——导致他们僵硬的创伤性事件,以及他们当时在自己的身心系统里察觉到的变化。
可以理解的是,这种讨论本身就会在参与者中激发应激唤醒——在很多群体中,有些人会在讨论过程中不自觉地进入僵硬状态。因此,这是一个具有挑战性的教学模块。指导老师必须引导整个团队降低他们的应激唤醒水平,同时为那些已经进入僵硬状态的人提供额外支持,而不是让他们引起别人的注意,因为这会让他们感到耻辱和羞愧。
在这个排的培训活动中,有关僵硬反应的讨论尤为热烈。很多士兵分享了他们自己的精彩故事。之前从未发言的海军士兵也急切地加入了讨论。这是全体学员第一次毫无反抗地积极参与培训活动。
然而,在讨论过程中,许多海军士兵出现了高度应激唤醒,包括胡里奥在内的三个人还进入了僵硬状态。我发现他们的眼睛变得呆滞而空洞,茫然地望着远处。他们的肩部前倾,身体一动不动。这些处于僵硬状态的海军士兵没有发言。
在这样的高度应激唤醒之下,我需要随机应变。通过使用一系列技术,我可以帮助大多数海军士兵降低他们的应激唤醒水平,让他们回到自己的耐受窗之内。在额外增加的课间休息中,我秘密地与胡里奥和另外两名进入僵硬状态的海军士兵一起练习;最终,他们也回到了自己的耐受窗之中。
尽管如此,我还是能看出胡里奥的僵硬反应很严重——他是第一个进入僵硬状态的人,随后恢复稳定的时间也更长。为此,我安排每一位海军士兵在接下来的几天内与我进行单独会面,谈谈他们到目前为止参加MMFT练习时的体验。现在,我可以轻松调整会面安排,以便在某天自然而然地与胡里奥会面,而不用单独挑他出来。
单独会面时,胡里奥告诉我在观看僵硬反应的视频时意外地发现自己出现了闪回,想起了童年时的一段经历,那是他很久以来都没有再想过的事情。在超过45分钟的时间里,胡里奥和我一同耐心地分析了整个过程,让他在耐受窗之内重新经历了相同的闪回过程,然后释放了压力。我用外部提示来引导他的注意力,我们一同帮助他的生存脑完成了一些必要的自下而上的恢复过程。我将在第三部分进一步探讨这个恢复过程。在本章,我想要探究的是胡里奥的思考脑和生存脑在闪回过程中究竟发生了什么。
胡里奥9岁时的某一天在外面和朋友及几个亲戚一起打球,这几个亲戚属于某个黑帮。突然,一辆卡车转过街角,在街道上加速行驶。当卡车里的人开枪时,胡里奥他们才发现枪手属于另一个敌对帮派。他感到一个表哥抓住了自己的胳膊,一路猛跑去寻找掩体。他看着自己的表哥向前跌倒了,但还是抓着胡里奥的胳膊。胡里奥感到自己也在跌倒,被他的表哥拽了下去。
突然间,一切都安静了下来。所有东西都失去了色彩。胡里奥仔细查看,发现自己的胳膊依然被表哥紧紧攥在手里。他想把胳膊收回来。他以慢动作向前跌倒,觉得自己会永远这么跌落下去。
在某个时刻,他感到自己好像又站了起来。他的肩膀剧痛,接着他看到了表哥的尸体。场面惨不忍睹,表哥的内脏被打出来了,浑身是血。
这时响起了一阵巨响,震耳欲聋。胡里奥听到了警笛声,还有人们的呼喊和哭泣声。他发现有人冲着他大喊大叫,但他却不知道他们在说什么。他觉得头晕,自己好像快要昏倒。
压力下的生存脑
正如应激唤醒与我们的神经系统和身体有关,它同样会影响我们的大脑,特别是学习和记忆功能。从进化的角度看,这是有道理的:学习和记住压力事件的能力对生存来说十分重要。思考脑和生存脑都有它们自己的学习和记忆模式,这使得它们能够发挥各自的功能。
然而,压力与创伤给思考脑和生存脑带去了不同的影响,同时还会对它们各自的学习和记忆过程产生连锁反应。本章探讨的正是这些差异。
压力和创伤源自生存脑的神经感知,这是一种无意识的评估过程,其功能是判断内外刺激究竟属于威胁、危险,还是机会、安全。生存脑中负责神经感知的部位被称为杏仁核,你可能还记得我们在第3章中说过,大脑的这个区域会随着不断的担忧而变厚。神经感知触发了一种基本上无意识的条件反射,而这种反应通常是趋利避害的。由于神经感知导致了条件反射,因此生存脑会进行“自下而上的处理”,也就是我们面对过往经历时的大多数下意识的情感和生理反应。经济学家丹尼尔·卡尼曼(Daniel Kahneman)将其描述为系统1思维,即“快思维”,因为它会在无须努力或自愿的情况下自动进行。[1]
由于生存脑并不具有语言功能,所以它不会通过思想或叙述与我们进行交流。[2]相反,它会产生情绪和生理感觉来触发这些条件反射式的防御措施和关系策略。由于生存脑不在意识范围之内,所以我们无法直接看到或了解这其中到底发生了什么。我们只能从身心系统的应激唤醒症状中看到它的作用。
为了实现神经感知功能,生存脑需要一个快速的学习和记忆系统。因此,它的学习系统是反射式的、无意识的、不自觉的且完全绕过了思考脑。
生存脑的学习系统被称为内隐学习(implicit learning)系统或系统1学习系统。内隐学习主要发生在杏仁核中,建立在总结以往神经感知经验的基础上。大多数这样的条件反射式学习都是在生存脑中进行处理和存储的。
内隐学习是由隐性(或非描述性)记忆(implicit or nondeclareative memory)支持的。[3]我们的每一次经历都在不断构建隐性记忆。当隐性记忆与掌握运动技能或基于身体的反应有关时,它被称为程序记忆(procedural memory)。例如,学习如何使用乐器、骑自行车、走路或跑步以及使用武器等都属于程序记忆。
当思考脑因为长期处于应激状态或遭遇极端压力而退化时,生存脑的学习和记忆功能会在任何应激唤醒水平下无意识地开启。此外,应激唤醒水平越高,生存脑的学习和记忆功能就越强。
在应激唤醒水平适度的情况下,杏仁核会与海马体一同制造外显记忆,其中的情感部分由杏仁核提供。[4]然而,在高度应激唤醒的情况下,杏仁核依然会提供记忆中的情感部分,而且强度更高,但海马体的功能却遭到了破坏。因此,在高度应激唤醒水平下,即便生存脑记下了很多东西,我们可能也无法强化有意识的记忆。因为海马体实际上可能会在高度应激唤醒水平下进入离线状态,所以对极具压力或创伤性的事件的有意识记忆通常都是不完整的、矛盾的或是支离破碎的。胡里奥的故事就是这种动因的例子之一,他讲述的部分细节特别生动,但其他部分却缺失了。然而,杏仁核可以从极端压力和创伤事件中学到和总结出的东西却是最多的。
让我们焦虑的大多数事情都是通过内隐学习造成条件反射的——这可能是因为我们的杏仁核无意识地将它与之前神经感知到的威胁联系在了一起,也可能是因为杏仁核根据它与此前神经感知到的威胁事件的相似性而将其泛化成了威胁。实际上,当隐性记忆和程序性记忆在危及生命的情况下被编码时,这些无意识的记忆就会变得更加持久,难以磨灭,同时也更容易将其他情形泛化为危机。
隐性记忆记住的不仅仅是事实或信息。[5]它记住的是神经系统的反应、身体感觉、肌肉和筋膜的张力、身体姿势、情绪以及防御模式下的运动模式。当我们在未来遇到相同威胁时,这些感觉和运动反应就会作为生存脑全部技能的一部分形成条件反射。这些条件反射可能会在我们没有意识到的情况下被唤起,就像那些在培训过程中出现僵硬反应的海军士兵一样。
在胡里奥的例子中,当他观看一只动物被捕食者按住的视频时,他的生存脑有可能将这一情形泛化成了他和表哥一同跌倒的经历。尽管那次跌倒的时候,胡里奥并没有被“按住”,但那种身陷囹圄、无助、无法摆脱表哥抓拽的感觉与之非常相似。胡里奥身体里的这种相似的感觉经自主神经系统的内脏传入系统传递到了他的生存脑中,这很可能是刺激他的生存脑开启僵硬反应的原因。
这里还有一个例子:我曾经培训过“山姆”,他是一位曾被派往阿富汗的海军陆战队士兵。几年后,在开始另一次征战之前,山姆每天很早就会醒来,他的心跳加速,呼吸急促,身体紧张得蜷缩成胎儿的姿势,他有一种很强的冲动想从床上跳起来。他不明白自己为什么会有这样的反应。
我向山姆解释说,即使他的思考脑无法理解,我们也必须相信他的生存脑这么做是有其重要原因的。下一次出现这种情况时,我建议他让自己的思考脑将挫败感放在一边,然后起床完成一项向下调节应激唤醒水平的练习(我将在第三部分教你进行这项练习)。
在连续一个星期每天早上都进行这样的练习之后,山姆的思考脑有了这样的结论:在他之前的征战经历中,很多时候他都是在天还没亮时被前方作战基地的炮火声惊醒的。可以理解的是,他的生存脑形成了强大的隐性记忆和程序记忆,也就是一醒来就要躲避迫击炮的袭击。现在,在知晓自己即将奔赴下一场征战的情况下,山姆的生存脑泛化了阿富汗战火下被反复唤醒的经历,给在家中的他制造了黎明前早起的焦虑。
压力下的思考脑
正如生存脑利用神经感知和内隐学习来保证我们的安全一样,思考脑也会对我们的生存发挥作用。思考脑保护我们的策略是分析、计划、审慎思考和决策。它进行的是“自上而下的处理方式”,也就是我们对自身经历的大多数自发和有意识的认知反应。[6]丹尼尔·卡尼曼将其描述为系统2思维,即“慢思维”。系统2确实是缓慢且费力的,它的特征是专注、有意识的思考和能动性。[7]
与生存脑的神经感知相似的是,思考脑负责的是执行功能(executive functioning),它主要发生在前额叶皮层。执行功能使我们能够集中注意力,保持专注,回忆与任务相关的信息,同时能够避免分心。[8]执行功能还可以让我们做出有意识的决策。我们利用执行功能“自上而下”地调节应激唤醒、冲动行为、渴望与情绪。因此,正如你能想到的那样,它也在控制“意志力”方面扮演了重要角色。
执行功能可能会在这几个方面遭到破坏。正如第3章解释的那样,我们会通过多任务处理破坏执行功能。我们还可以通过酒精、毒品来破坏它,而这正是我们在醉酒时很难控制自己的原因。此外,压力也会对它造成损害。
执行功能好似信贷银行,我们可以通过两种方式掏空它。[9]一方面,我们可以执行所谓的“冷”认知任务,即需要细致关注和聚焦的心理任务,例如,阅读大量文本,撰写报告或完成繁重的计算等。另一方面,我们还可以执行“热”调节任务,即为了抑制欲望、重塑或隔离消极情绪,以及抑制应激唤醒而进行的有意识的、自上而下的努力。
每当我们通过“冷”任务或“热”任务消耗执行功能时,银行中用于支付两种任务成本的信用额度就会减少。这也就解释如下两种情形:在我们审核了一整天烦冗的财务文件,也就是完成了“冷”任务之后,当有人挡了我们回家的路时,我们很有可能会陷入沮丧情绪,与其发生冲突;或当我们回到家时,我们很有可能会打破自己的节食计划。留给“热”调节任务的额度所剩无几。
相反,当我们在一段重要的人际关系中长期处于紧张状态或是在工作中遭遇长期歧视(这两种状态都需要持续的“热”调节)时,我们可能必须将一段文字读上7遍才能最终理解它的意思,这是因为没什么可以留给理解这一“冷”任务的了。
无论执行功能遭到了怎样的破坏或消耗,在这种状态下,压力和情绪更有可能驱动我们的决策。[10]我们的思考脑思考问题的方法会因压力和情绪而产生偏差。我们也更有可能做出习惯性的、冲动的、反应性的暴力或不道德行为。
为了支持执行功能,思考脑需要一个学习和记忆系统,这个系统强调的是置于时空背景之下的语境信息。思考脑的学习系统进行的是自主学习或系统2式学习。自主学习主要发生在海马体中。
自主学习是由显性记忆或陈述性记忆(explicit or declarative memory)支持的,比如对事件、事实、容貌、字词或信息的记忆等。[11]和隐性记忆不同,我们是有意识地获取显性记忆的,例如,回忆自己的人生故事或试图将新信息加入我们的“知识库”。在我们两岁之前,海马体中的神经纤维不会形成脂肪鞘(这个过程名为髓鞘化),更不会通过它来导电,因此我们很少会形成关于幼年时光的显性记忆。
显性记忆受我们的智力和其他个人差异影响,但正如胡里奥的故事反映的那样,它也深受我们应激唤醒水平的影响。[12]
轻度至中度的应激唤醒水平能够增强短期内的显性记忆和自主学习水平。[13]在轻度到中度的刺激之下,高皮质醇和高血糖水平意味着海马体能够获得现成的能量,而这可以让我们集中注意力,有助于显性记忆的形成、存储和检索。实际上,大脑对葡萄糖是非常贪婪的,虽然它仅占我们体重的3%,却消耗了人体中20%的循环葡萄糖,其中海马体是最贪爱葡萄糖的部位之一。[14]
这就是喝咖啡会提高我们的注意力的原因,因为它引起了我们皮质醇水平的飙升。这也是低血糖在我们感到饥饿的时候会导致执行功能和显性记忆下降,同时令我们更加易怒的原因。一项研究调查了以色列假释裁决委员会的法官和他们的用餐休息时间。[15]研究发现,距离法官们上一餐的时间越长,他们越有可能否决假释。在感到“饥饿”时,他们调节易怒情绪的能力就会减弱,而在法庭上带着清晰的思路和同情心来评判案件细节的能力也会减弱。
相对于轻度至中度的应激唤醒水平,执行功能和显性记忆功能可能会在慢性压力或高压下受损。[16]为什么会这样呢?过高或长期升高的皮质醇水平会导致海马体内的神经元失去浓密的树突(连接相邻神经元的分支),从而导致神经网络萎缩。随着长期应激唤醒的持续,原有神经元死亡,新的神经元停止生长,海马体体积缩小。实际上,在长期或极端压力之下,大脑将暂停海马体中有益的神经重塑和神经发生。
我们常常将大脑的这些变化视为记忆问题。我们可能难以集中注意力、学习新知识、做出规划和执行任务。我们可能在屏蔽干扰方面有困难。每一个这样的征兆都表明我们的执行功能和显性记忆能力遭到了损耗和破坏。
还记得第3章针对派驻伊拉克的士兵的研究吗?[17]与那些没有被部署到战场的士兵相比,这些士兵的反应更快,这标志着他们的生存脑在高压作战状态下超时工作了。然而,在回家两个月后,生存脑的这种转变的代价显现了出来,表现为思考脑的专注力、执行功能和显性记忆衰退。
针对高压工作的研究包括内科住院医师项目、执法工作、消防人员灭火演习以及军事部署和野战演习等,表明个人在这些环境下往往会经历更多焦虑和痛苦。[18]他们还会出现更多认知能力退化的症状,包括精神分裂、思维混乱、解决问题能力不足、注意力分散、视觉模式识别困难以及工作记忆衰退等。
因为应激唤醒也伴随着象征威胁,比如对未来负面事件的预想或过去创伤记忆的“点燃”,所以,即便我们没有遭遇直接的身体伤害,思考脑的功能也还是会衰退。[19]
在长期或极端压力的作用下,任何人都可能出现执行功能和显性记忆衰退,哪怕他们从事的并不是对生命有威胁的职业。这些衰退可能是在长期睡眠不足之后或在经历换工作、搬家、结婚、生孩子等重大人生转变期间发生的。
衰老同样会加剧这些影响。[20]有关老年健康的前瞻性研究表明,那些皮质醇水平在多年研究期间有所增加的老年人的记忆衰退程度最高,海马体体积的损失也最多。
实证研究表明,对遭遇多种不同情况的人群来说,过高的压力或皮质醇水平的升高与海马体体积的缩小和记忆问题有关。例如,洲际航班乘务员要长时间工作,同时要切换时区,经历慢性时差反应,但却无法在下一次起航前得到充分恢复[21];长期使用处方类固醇(比如氢化可的松乳膏、可的松注射液、口服或吸入类固醇等)的人[22];长期患有重度抑郁症的人[23];以及在反复遭遇创伤后患有创伤后应激障碍的人,比如在童年反复遭遇虐待或长期经历战争的幸存者[24]。在这些情况下,压力水平越高,持续时间越长,记忆问题就越严重,海马体的体积也更小。
耐受窗如何影响思考脑和生存脑之间的关系
应激唤醒和人的表现水平(包括思考脑的功能)之间的关系是一个倒U型的耶基斯-多德森曲线(Yerkes-Dodson Curve)(见图5-1)。在应激唤醒水平较低的情况下,我们可能没有足够的应激唤醒水平保持警醒,也没有足够动力来有效完成任务。实际上,良性压力(eustress)——采用了希腊语中含义为“良性”的前缀“eu”——能够提供注意力和能量。思考脑的功能会因良性压力而增强,因为轻度或中度压力会使人体循环中的葡萄糖和皮质醇水平升高,让我们集中注意力,记住与任务相关的信息。
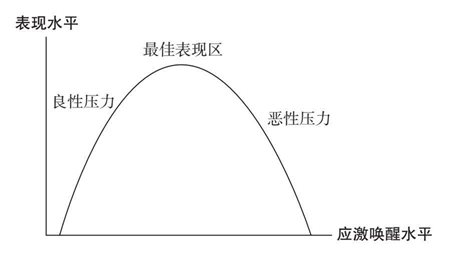
图5-1 耶基斯-多德森曲线
1908年,心理学家罗伯特·M. 耶基斯(Robert M. Yerkes)和约翰·D. 多德森(John D. Dodson)首次提出了应激唤醒水平和表现水平的倒U型关系,也就是今天所说的耶基斯-多德森曲线。当一个人接近中等应激唤醒水平时,这个人在执行任务时所表现出的水平就会达到最佳,但超过这一应激唤醒水平之后便会稳步下降,直至进入彻底僵硬状态。良性压力(“好”的压力)指的是低度应激唤醒水平,而恶性压力指的是高度应激唤醒水平。
相反,在高度应激唤醒水平下,我们的表现会稳步下降。不出意料的是,最终的极端表现就是僵硬。在曲线的恶性压力一侧,思考脑功能受损则令人难以集中注意力,排除干扰,记住与任务相关的信息,调节自己的压力和负面情绪,也无法做出有效决策。
因此,最佳表现、自主学习以及有效决策最有可能在中度应激唤醒水平下产生,在这种情况下,应激唤醒水平足够让我们保持警觉和专注,但不足以进入恶性压力区。[25]考虑到这一点,我们神经生物学中针对应激唤醒的耐受窗实际上就是这样一个窗口:我们在其中能够上下调节压力水平,让我们在时间的推移下保持在中度应激唤醒的最佳表现区之中。[26]
在这个窗口中,思考脑和生存脑作为联盟一同运作,而它们也完全发挥了各自的功能。在我们的耐受窗中,我们更有可能进行准确的神经感知,也就是说,如果我们感知到危险,那么客观情况一定真的十分危急。[27]我们更有可能感知到相关的内部和外部线索,然后去理解它们,对它们进行客观评估,并将所有这些新数据与我们已知的影响当前状况的信息整合起来。我们更有可能对我们所有选择的成本和收益进行思考与评估。接下来,我们就更有可能做出符合我们价值观和目标的选择。最后,我们更有可能有意识地评估这一决定的结果,从而能够适应未来并从中有所收获。因此,我们的耐受窗越大,我们越有可能保持准确的神经感知,有效整合思考脑和生存脑的活动,哪怕是在高度应激唤醒和情绪紧张的状态下。
要想达到最佳表现状态,我们必须有能力调节自己的应激唤醒水平,以适应手头的任务。不同的任务需要不同的应激唤醒水平。[28]例如,当我们尝试新事物时,我们更有可能在较低的应激唤醒水平上取得成功。正如第4章解释的那样,这是因为生存脑认为新事物更具威胁性。相反,单调或熟悉的任务可能要求更高的应激唤醒水平来创造专注力和动力。实际上这正是人们拖延那些让人不愉快的任务的原因之一:随着最后期限的临近,他们的应激唤醒水平增强,最终产生了足够的压力去推动他们处理这项工作。
相反,当我们的应激唤醒水平和紧张情绪超出了耐受窗,我们更有可能产生错误的神经感知。在耐受窗之外,我们更有可能在情况没那么糟糕的时候感知到危险,继而激发保护性行为作为响应。抑或在情况实际相当危险时,我们可能会感知到安全和机遇,而这将导致我们陷入危险的境地。因此,在应对手头的情况时,我们的应激唤醒水平可能要么过高,要么过低。
在我们的耐受窗之外,生存脑更有可能驱动信息搜索、评估及决策制定等行为,思考脑则同时以退化或受损的方式进行运作。[29]高压往往会缩小我们的感知范围,因此我们会无意识地将注意力和优先级放在眼前的事情上,而不是那些对我们的长期成功而言真正重要的地方。我们总是会带着偏见去理解事物的意义。我们强调的是自己心理上认为重要的信息,专注于压力源和压力本身。[30]在这个过程中,我们失去了洞察力。当我们感到有压力时,我们搜集的信息通常也更少。[31]我们更有可能从少量信息中得出结论,而这些结论往往是断章取义的。此外,我们也会对负面信息产生偏见,因为负面信息更有可能引起我们的注意,得到更及时的处理,回忆起来也更加清晰。[32]这在进化上是说得通的。如果我们旧石器时代的祖先能够从负面的压力事件中快速学习经验并且记住它们,那么他们更有可能生存下来。
感知范围缩小和僵化的情况同样会出现在决策制定过程中。[33]我们更有可能接受第一个可行性选择,而不会对所有选择加以考虑。我们更多地依赖刻板的印象、过度简化的假设和过往的经验,而没有清晰地看到当前局势的独特性。我们分析复杂情况、战略互动和决策后果的能力下降,同时也更容易犯错。
在耐受窗之外,思考脑和生存脑形成了一种对抗关系,这种关系表现在三个方面。第一,思考脑的功能衰退可能会表现为不准确的态势感知、记忆力问题、注意力不集中、焦虑的规划思路或防卫性思维等。
第二,我们可能会受到生存脑劫持。在这种情况下,情绪和应激唤醒会让我们的感知产生偏差,抓住我们的大部分注意力,驱动我们的决策与行为。这时,生存脑和神经系统可能会进入默认程序,即便这并不是当前情况下最合适的选择。当这个程序下意识地进行时,我们的思考脑可能会试图将其归咎于他人或我们自己。
第三,我们可能会受思考脑控制,也就是“活在我们自己的思维里”,而与生存脑的信号(包括我们的情绪和生理感觉)断开联系。几乎可以肯定的是,当陷入压抑、否认或区隔的状态,或者当咬咬牙,继续前进时,我们就会进入思考脑控制状态。
当然,在不同的情况下,我们可能会经历这三种对抗关系。在生存脑完全有效恢复之前,这些模式都将继续下去,同时进一步缩小我们的耐受窗。随着时间的推移,这些动因将会造成非稳态负荷,导致我们产生失调症状。
创伤下的生存脑
创伤和慢性压力用来缩小耐受窗的途径是相互关联却又各不相同的。创伤与更多动因有关,这些动因将思考脑和生存脑之间的关系进一步复杂化了。其中最重要的一种外加的动因在于,生存脑的隐性记忆系统在创伤期间遭到破坏。它认为我们无力、无助,缺乏对事物的掌控。此后,这种无助感便深深地植入隐性学习过程之中。我们是否真正幸存下来并不重要,因为生存脑认为我们不能成功保护自己。
例如,我们可能会身陷囹圄或身体受限,无法反击或逃跑,或者被人制服。我们可能会经历交通事故、自然灾害、恐怖袭击或大规模枪击,这些事件完全超出了我们的控制。在更微妙的情况下,我们可能会在工作中遭受歧视或骚扰之类的窘境,而为自己或他人辩护可能会让我们失去工作,因此我们保持沉默或默许了这些行为。当生存脑根据这些创伤事件对我们的隐性记忆进行编码时,它会无意识地将高度应激唤醒水平和无力、无助及失去掌控感联系起来。[34]
在胡里奥的闪回中,他无法摆脱表哥的抓拽,无法阻止自己跌倒,也无法阻止表哥的死亡。他的生存脑可能得出了这样的结论:他在感到“受约束”时无法成功保护自己。
隐性记忆系统将来自创伤事件和“成功”(完成)防御事件的记忆区分开来。在未能成功防御和恢复的情况下,创伤事件中激活的压力就无法得到释放。这样一来,创伤记忆就会以一种确保其能持续下去的方式得到编码。
因此,生存脑认为创伤事件永远不会结束。关键在于,直到生存脑、神经系统及身体得到机会去完成不完整的防御策略并且释放相关压力,生存脑才能继续感知进行中的事件。[35]生存脑还会认为自己无力成功抵御进行中的威胁,而这又增加了再次陷入僵硬状态的可能性,就像胡里奥在观看动物视频时的表现一样。
经历创伤事件之后,生存脑的隐性记忆遭到破坏会带来这样三个后果。第一,由于生存脑认为创伤事件还在进行之中,所以它无法完成完整的处理过程,也无法从该事件中学到什么。与此同时,它还将继续严重依赖于它在最初的创伤事件中使用的不成功的防御策略。这就好像生存脑下意识地认为:“如果在当前情况下采用相同的防御策略,也许这一次它就奏效了,最后我也能成功保护我自己。”
这意味着受到创伤的生存脑失去了分辨当下和过去的能力,因此无法进行学习和调适。在没有完全恢复的情况下,生存脑在时间的流转之中依然处于僵硬状态,好像回到了最初创伤事件发生的时候。[36]可以理解的是,这种动因助长了创伤重演,它将无意识地再造一些情境与关系,而它们在某些方面是与早期创伤事件相呼应的。这也是为什么创伤幸存者,特别是那些在童年遭遇创伤的人很容易在今天的危险状况下做出与当前实际情况不相关,而且常常适得其反或有害的大脑反应。面对当下的危险,生存脑唯一可用的习得反应是重复最初创伤事件发生时的默认条件反射行为。
创伤事件中编码不完整或不成功的防御策略,被生存脑当作了和最初创伤事件相似的情形下的全新通用默认策略。在胡里奥的例子中,当他看到视频中的动物“被束缚”时,他的体内激发出了当他跌倒时被表哥紧紧“束缚”的相同感受。这种相似性足以让他的生存脑在培训中再度触发默认的僵硬程序。
在环境改变很久之后,受到创伤的生存脑依然准备使用它的默认策略。[37]这些默认策略取代了有关当前情况的信息,占据了优先地位,哪怕有其他更加合适的策略。生存脑将继续依赖这种默认程序,直到它能完全恢复。只有在那时,生存脑最终才能认识到创伤事件真的已经过去,它的身心系统现在是安全的。
隐性记忆系统受损的第二个后果是,生存脑对有关最初创伤的任何线索都变得极度敏感。
生存脑在创伤事件中捕捉到的大部分信息都不在思考脑的意识之中,而是被当作记忆胶囊的一部分存储在隐性记忆系统之中。记忆胶囊包含感觉输入(视觉、听觉、嗅觉、味觉和触觉)、身体姿势、身体动作、情绪及生理感觉,特别是和交感神经系统与背侧副交感神经系统应激唤醒有关的感觉。[38]记忆胶囊可能还包括感知改变,例如,感到自己好像正从天花板上看着自己或感到时间慢下来等。
每个记忆胶囊的强度取决于创伤事件的强度,以及该事件是否在一段时间内重复发生。对我们在创伤事件中经历的应激唤醒和情绪强度来说,如果我们还没有在彻底的恢复中释放它们,它们就会像“胶水”一样将记忆胶囊的各个组成部分黏合在一起。在完全恢复之前,记忆胶囊仍处于活跃状态,很容易被触发。
当生存脑感知到今天类似于我们未得到解决的记忆胶囊一部分的线索时,这个线索就可能触发记忆胶囊。反过来,身心系统的反应就好像这种创伤现在再度发生,而且恐惧和无助程度也和最初一样。这就好似我们可以通过许多各不相同的无意识之门进入一个房间,房间里尽是没有得到解决的创伤。
这也解释了退伍军人或枪击暴力事件幸存者为什么可能会在听到汽车回火声时经历闪回。实际上,他们的杏仁核泛化了回火的声音,将其与存储在记忆胶囊里的枪击声联系在了一起。
重要的是,由于记忆胶囊存储在生存脑的隐性记忆系统之中,因此思考脑无法理解记忆胶囊触发的机制和原因。事实上,思考脑甚至可能没有意识到记忆胶囊的触发。这因为记忆胶囊是在高度应激唤醒的前提下得到存储的,也就是杏仁核记忆最好的时候,而思考脑的显性记忆可能已经中断或离线。[39]这就是为什么创伤事件的有意识记忆可能是模糊的、支离破碎的和自相矛盾的。某些细节可能在有意识的记忆中烙下水晶般清晰的印记,而其他细节则完全丢失了。
隐性记忆系统受损的第三个后果是,每当记忆胶囊被触发时,生存脑和身体就会产生症状,表明生存脑相信创伤事件仍在继续。[40]这些症状包括闪回、噩梦、侵入性思维、担忧、强迫性思考,以及与当前情形不相称的应激唤醒下的身体症状等。通过产生这些症状,生存脑试图完成最初的创伤事件,重新恢复非稳态负荷,但没有成功。这些症状突出表现了生存脑和身体并不理解那件事情实际上已经过去。相反,它们继续调动应激唤醒来应对这种被误解的持续威胁。
第三个后果的代表是点燃效应(kindling),也就是过往的良性事件触发应激唤醒的情况。点燃效应发生在未解的记忆胶囊被反复触发的情况下,比如闪回,而每一次触发都会加剧症状。最终,应激唤醒的内部征兆(比如呼吸短促、肌肉紧张、恶心及摆出某些姿势等)可能会成为完全独立于任何外部征兆的应激唤醒源。这就是这种状况被称为“点燃效应”的原因,因为这种唤醒源就像是用来生火的易燃枝条。
点燃效应每发生一次都会“变本加厉”地增加身心系统对过去征兆的敏感度。[41]随着时间的推移,身心系统可能会进行自我激活,从而变得与当前环境越来越不协调。
实际上,受到创伤的生存脑会将它在身心系统内部感知到的进行中的危险投射到外部环境中去,这些危险与未解的记忆胶囊相关。它陷入了过去,将没有得到解决的危险记忆胶囊叠加在预设的情形之上,哪怕这种情形毫无危险之处。这种扭曲是生存脑劫持的极端现象。在这种情况下,未解的记忆胶囊无意识地扭曲了受到创伤之人的观点,吸引并捕获了他们的注意力,促使他们采取默认的防御措施和关系策略,同时影响了他们的人际关系和决策制定。[42]
随着时间的推移,生存脑会对这些内部压力源(比如身体感觉、疼痛、令人痛苦的念头以及与最初创伤有关的情绪)产生这样的感知,认为它们变得越来越具有威胁性和挑战性。这就是为什么早期创伤事件造成的症状会随着时间的推移而加剧,就好像它们“有了生命一样”。而它们通过点燃效应的确也做到了。[43]
创伤下的思考脑
受到创伤的人还会让情况变得更加复杂。这是因为在经历创伤之后,大脑负责认知、情绪和感觉运动水平的不同部分之间信息处理的整合过程常常会遭到损害。[44]而这放大了思考脑和生存脑之间的对立关系,同时为彻底恢复增加了额外的障碍。
人类从生存脑(杏仁核)到思考脑(前额叶皮层)的神经回路比反向回路更多,规模也更大。[45]这在进化上是说得通的,因为对威胁的快速评估和反应增加了我们的生存机会。此外,在慢性压力或创伤性压力之下,源自生存脑的神经回路会得到更多锻炼,而源自思考脑的神经回路则可能会受损或退化。总之,这两种不平衡有助于解释为什么当我们身处耐受窗之外时可能会发生生存脑劫持现象。
更重要的是,这两种不平衡也说明了为什么思考脑很难纠正生存脑受损的隐性记忆系统。这个被称为恐惧消退或创伤消退的过程,依赖于我们在慢性压力或创伤性压力中退化的思考脑回路。这种消退实际上是一种新记忆的生成,而非对我们现有记忆的消除。
在经历创伤事件之后,思考脑认为该事件已经过去,而我们生存了下来。由于思考脑知道事情已经过去,所以它通常会使用其思考、分析、计划、深思熟虑和决策的保护机制来保障我们未来的安全。它可能会对该事件进行分析,从中学习并为其赋予一定意义,它会谴责他人,进行自我批判,思考我们如何引发了该事件或制订计划来防止这样的事情再度发生。
换句话说,对思考脑而言,创伤已经过去,它的行动是利用其工具,继续前进。
思考脑的行动过程与生存脑的创伤后认知和保护计划完全相反。毕竟,生存脑认为创伤事件还在持续。因此,当生存脑感知到与未解记忆胶囊有关的征兆(包括通过点燃效应产生的应激唤醒的身体感觉)时,它会继续感知危险,调动应激机制。然而,矛盾的是,由于生存脑并不能支持思考脑认为创伤已经过去的“理性”认知,它们各自认知之间的不一致实际上可能会导致生存脑产生更加不安全的感觉。[46]
这就好像生存脑在说:“我发现身体里出现了种种应激唤醒现象。既然身体已被激活,那么一定有威胁,对吗?但我还没有看到威胁。因此,一定是有什么事蒙蔽了我,而危险可能比我想象的还要严重。”这种动因有助于解释为什么创伤幸存者常常觉得自己如履薄冰、提心吊胆。这正是他们的生存脑不自觉地想要将相对安全的外部环境和内心的危险感觉联系在一起的表现。
因此,这一过程通常会引发恶性循环也就不足为奇了,这会使受到创伤的生存脑将中性乃至积极的刺激视为威胁。实际上,受到创伤的幸存者可能会渐渐对体内各种形式的应激反应产生怀疑,哪怕是运动、跳舞或做爱时产生的愉悦的兴奋感。例如,在愉悦的事件中,思考脑可能会享受正在发生的事情,但同时受到创伤和过度警觉的生存脑则专注于寻找威胁来“解释”这种兴奋感。
遭遇创伤后,思考脑和生存脑对当前情况的理解存在巨大差异,生存脑会继续产生危险的感觉也就不难理解了。然而,由于思考脑认为创伤事件已经结束,所以它通常不理解身心系统为何会如此运作。
思考脑可能会尝试用对抗性的思考脑习惯去分析我们的行为和症状。例如,它可能会产生批判性的想法和比较性的想法,例如,“我现在应该结束这一切,我到底怎么了?其他人可能做得更糟糕。”思考脑也有可能会进行自我评判,产生内疚和羞愧的情绪。它可能会有焦虑的想法,担心这些症状永远不会消失或进一步恶化。所有这些对抗性的思考脑习惯只会加剧应激唤醒。
为了回应这些“分析”,思考脑可能会认为应激唤醒和失调症状是亟待解决的“问题”。这么做可能会导致思考脑控制,例如,对问题进行压制和隔离,咬咬牙,继续前进,奋勇向前。当我们生活在“自己的思想中”而脱离了情感、直觉和躯体时,思考脑控制的状况也会显现。
比如认知行为疗法、积极心理学、暴露疗法、其他谈话疗法、认知再评价或目标设定法等治疗手段可能也会在不经意间激励思考脑控制。思考脑主导的手段旨在通过强化自我,抑制应激唤醒,抑或让我们失去对未解记忆胶囊线索的敏感性来实现自上而下的自我调节,因为它认为应激唤醒才是需要解决的“问题”。
然而,如果这些手段没有与自下而上的处理过程相结合,那么在我们的耐受窗之内,生存脑便无法对受损的记忆系统进行更新。因此,仅仅使用这些技术可能会使受创伤者将他们的应激唤醒水平严格限制在他们狭窄的耐受窗中,从而避免触发他们未解决的记忆胶囊。[47]然而,与此同时,他们还是处在隔离状态,与他们自身相信创伤还在继续的那一部分相互分离。反过来,生存脑和身体继续用未解的记忆胶囊覆盖当前状态,它们无意识地依赖默认创伤防御程序,并通过点燃效应制造越来越严重的失调症状。
因此,正如创伤治疗医师帕特·奥格登和她的同事解释的那样,尽管思考脑主导的手段“为过度兴奋提供了有效管理,显着缓解了症状,但它们可能无法彻底解决这一问题”。[48]
可以肯定的是,在特定情况下,思考脑可以在短时间内非常熟练地发挥控制作用。尽管如此,这些手段本身并不能促进拓宽耐受窗所需的彻底恢复。实际上,采用这些手段时,思考脑可能会感到更有控制力,而生存脑对无助和缺乏控制力的感觉可能还将持续。因此,长期依赖这些方法可能会使未解的记忆胶囊永续下去,同时增加非稳态负荷。
最终,思考脑的控制可能也会积极抑制生存脑对恢复的尝试。我们大多数人从未被教导过如何在身心系统中释放应激唤醒。(你将在第三部分了解到这方面的内容。)因此,当这些感觉和习惯自发产生时,我们可能会推翻和抑制它们,特别是当它们落入思考脑的叙事和文化范式时,比如“男孩不哭”。
思考脑的控制作用越发极端,生存脑的应激唤醒水平就越高,它可能会向思考脑传递这样的信息:“我现在不安全!”为了让它的信息能够传递过去,它采取了生存脑劫持手段(比如闪回、噩梦以及呕吐在键盘上),这也是失调的一种表现。作为回应,思考脑会加倍抑制、隔离,让人咬紧牙关,继续前进,无视生存脑发出的信号。实际上,它会产生这样的想法:“我到底怎么了?这些都是过去的事了。我现在应该已经跨过这个坎了。”
每当思考脑阻碍了生存脑试图恢复的努力,它就会加剧我们的失调症状。更重要的是,它还会进一步肯定和重建应激唤醒与生存脑无助感之间的联系,这种神经生物学上的重要耦合正是创伤性压力的特点。实际上,生存脑会将“我无法成功保护自己”的创伤思维泛化为“我无法成功恢复”的想法。这种对彻底康复的无助感进一步锁定了创伤模式,造成恶性循环。
对经历过创伤的人来说,这是一个难解的死结:体内的应激反应、未解的记忆胶囊、在创伤事件中采用的不完善或不成功的默认防御手段和关系策略以及当前无法彻底恢复的习得性无助。在解开这个死结之前,生存脑和身体会继续认为创伤事件仍在进行之中,因此它们会依赖创伤中的默认程序。
然而,由于思考脑和生存脑对之前的创伤有着相当不同的理解,所以思考脑常常会在不知不觉中成为从未实现过的全面恢复的主要障碍。相反,为了应对日益严重的失调症状,大多数受过创伤的人都会采用一系列被社会接受的行为来应对——然而可悲的是,这么做只会进一步缩小耐受窗。
在开始下一章之前,我想鼓励你带着不加评判的好奇心,回忆你经历过的思考脑控制,比如隔离或生存脑劫持,如闪回或点燃效应。在你的笔记中,你可以问问自己,这些心理习惯是否是由特定的情况、人际关系或记忆所触发的。当这些习惯被触发时,你通常又是如何应对它们的?
治愈思考脑和生存脑之间的对立关系的第一步是意识到它的存在。在理解了它的神经生物学动因之后,随着时间的推移,我们便可以促进生存脑实现彻底的恢复,同时消除它与思考脑之间的对立关系。
[1]有关系统1思维的特征,可参考: Daniel Kahneman, Thinking, Fast and Slow (New York: Macmillan, 2011), 105。
[2]Pat Ogden, Kekuni Minton, and Clare Pain, Trauma and the Body: A Sensorimotor Approach to Psychotherapy (New York: Norton, 2006), chap. 1;Robert C. Scaer, The Trauma Spectrum: Hidden Wounds and Human Resiliency(New York: Norton, 2005), chap. 2; Stephen W. Porges, The Polyvagal Theory:Neurophysiological Foundations of Emotions, Attachment, Communication,and Self-Regulation (New York: Norton, 2011), chaps. 1, 3.
[3]Scaer, The Trauma Spectrum, 40-42.
[4]Scaer, The Trauma Spectrum, 40-42; Robert M. Sapolsky, Why Zebras Don't Get Ulcers, 3rd ed. (New York: Holt, 2004), chap. 15; J. Douglas Bremner,Does Stress Damage the Brain?: Understanding Trauma-Related Disorders from a Mind-Body Perspective (New York: Norton, 2005), chap. 4; Bruce S. McEwen and Elizabeth Norton Lasley, The End of Stress as We Know It(Washington, D.C.: Joseph Henry, 2002), 108-110.
[5]Sapolsky, Why Zebras Don't Get Ulcers, 320-322; Scaer, The Trauma Spectrum, 40-42; J. LeDoux, “The Emotional Brain, Fear, and the Amygdala,”Cellular and Molecular Neurobiology 23, no. 4-5 (2003): 727-738; McEwen and Lasley, The End of Stress as We Know It, chap. 7; Bremner, Does Stress Damage the Brain?, chap. 4.
[6]Paul D. MacLean, The Triune Brain in Evolution: Role in Paleocerebral Functions (New York: Plenum, 1990); Ogden et al., Trauma and the Body,chap. 1.
[7]Kahneman, Thinking, Fast and Slow, 21; J. Evans, “In Two Minds: DualProcess Accounts of Reasoning,” Trends in Cognitive Sciences 7, no. 10 (2003):454-459.
[8]K. N. Ochsner and J. J. Gross, “The Cognitive Control of Emotion,” Trends in Cognitive Sciences 9, no. 5 (2005): 242-249; J. М. Hinson, T. L. Jameson,and P. Whitney, “Impulsive Decision Making and Working Memory,” Journal of Experimental Psychology: Learning, Memory, and Cognition 29, no.2 (2003): 298-306; W. Hofmann, B. J. Schmeichel, and A. D. Baddeley,“Executive Functions and Self-Regulation,” Trends in Cognitive Sciences 16,no. 3 (2012); 174-180; W. Hofmann et al, “Working Memory Capacity and Self-Regulatory Behavior: Toward an Individual Differences Perspective on Behavior Determination by Automatic versus Controlled Processes,” Journal of Personality and Social Psychology 95, no. 4 (2008) 962-977; M. L. Pe, F.Raes, and P. Kuppens, “The Cognitive Building Blocks of Emotior Regulation:Ability to Update Working Memory Moderates the Efficacy of Rumination and Reappraisal on Emotion,” PLOS One 8, no. 7 (2013): e69071; T. F. Heatherton and D. D. Wagner, “Cognitive Neuroscience of Self-Regulation Failure” Trends in Cognitive Sciences 15, no. 3 (2011): 132-139.
[9]Hofmann et al. “Executive Functions and Self-Regulation.”有关睡眠不足对执行功能的影响可参考:L. K. Barger et al., “Neurobehavioral, Health,and Safety Consequences Associated with Shift Work in Safety-Sensitive Professions,” Curren Neurology and Neuroscience Reports 9, no. 2 (2009):155-164。
[10]Heatherton and Wagner, “Cognitive Neuroscience of Self-Regulation Failure”;Hofmann et al., “Executive Functions and Self-Regulation.”
[11]Scaer, The Trauma Spectrum, 38-39; Bremner, Does Stress Damage the Brain?, 45-47.
[12]Scaer, The Trauma Spectrum, 38; Pe et al., “The Cognitive Building Blocks of Emotion Regulation”, Hofmann et al., “Working Memory Capacity and SelfRegulatory Behavior”.
[13]Sapolsky, Why Zebras Don't Get Ulcers, chap. 10.
[14]McEwen and Lasley, The End of Stress as We Know It, 130; E. R. De Kloet et al., “Brain Corticosteroid Receptor Balance in Health and Disease,” Endocrine Review 19, no. 3 (1998): 269-301.
[15]S. Danziger, J. Levav, and L. Avnaim-Pesso, “Extraneous Factors in Judicial Decisions,” Proceedings of the National Academy of Sciences 108, no. 17 (2011): 6889-6892. 另见M. T. Gailliot and R. F. Baumeister,“The Physiology of Willpower: Linking Blood Glucose to Self-Control,”Personality and Social Psychology Review 11 (2007): 303-327; J. M.Tyler and K. C. Burns, “After Depletion: The Replenishment of the Self's Regulatory Resources,” Self and Identity 7 (2008): 305-321.
[16]有关这一动因的精彩评述,可参考:McEwen and Lasley, The End of Stress as We Know It, chap. 7; Bremner, Does Stress Damage the Brain?,chap. 4; Sapolsky, Why Zebras Don't Get Ulcers, chap. 10。
[17]J. J. Vasterling et al., “Neuropsychological Outcomes of Army Personnel Following Deployment to the Iraq War,” Journal of the American Medical Association 296, no. 5 (2006): 519-529.
[18]B. Vila, G. B. Morrison, and D. J. Kenney, “Improving Shift Schedule and Work-Hour Policies and Practices to Increase Police Officer Performance,Health, and Safety,” Police Quarterly 5, no. 1 (2002): 4-24; A. Gohar et al., “Working Memory Capacity Is Decreased in Sleep-Deprived Internal Medicine Residents,” Journal of Clinical Sleep Medicine 5, no. 3 (2009):191-197; Barger et al., “Neurobehavioral, Health, and Safety Consequences”;M. R. Baumann, C. L. Gohm, and B. L. Bonner, “Phased Training for HighReliability Occupations: Live-Fire Exercises for Civilian Firefighters,”Human Factors 53, no. 5 (2011): 548-557; H. R. Lieberman et al., “Severe Decrements in Cognition Function and Mood Induced by Sleep Loss, Heat,Dehydration, and Undernutrition during Simulated Combat,” Biological Psychiatry 57, no. 4 (2005): 422-429; H. R. Lieberman et al., “Effects of Caffeine, Sleep Loss, and Stress on Cognitive Performance and Mood during U.S. Navy Seal Training,” Psychopharmacology 164, no. 3 (2002): 250261; Jennifer Kavanagh, Stress and Performance: A Review of the Literature and Its Applicability to the Military (Arlington, VA: RAND, 2005); C. A.Morgan et al., “Stress-Induced Deficits in Working Memory and VisuoConstructive Abilities in Special Operations Soldiers,” Biological Psychiatry 60, no. 7 (2006): 722-729; C. A. Morgan et al., “Accuracy of Eyewitness Memory for Persons Encountered during Exposure to Highly Intense Stress,”International Journal of Law and Psychiatry 27, no. 3 (2004): 265-279;C. A. Morgan et al.,“Symptoms of Dissociation in Humans Experiencing Acute, Uncontrollable Stress: A Prospective Investigation,” American Journal of Psychiatry 158, no. 8 (2001): 1239-1247; C. A. Morgan et al., “Neuropeptide-Y, Cortisol, and Subjective Distress in Humans Exposed to Acute Stress: Replication and Extension of Previous Report,” Biological Psychiatry 52, no. 2 (2002): 136-142; C. A. Morgan et al., “Relationships among Plasma Dehydroepiandrosterone Sulfate and Cortisol Levels,Symptoms of Dissociation, and Objective Performance in Humans Exposed to Acute Stress,” Archives of General Psychiatry 61, no. 8 (2004): 819-825;C.A. Morgan et al., “Relationship among Plasma Cortisol, Catecholamines,Neuropeptide Y, and Human Performance during Exposure to Uncontrollable Stress,” Psychosomatic Medicine 63, no. 3 (2001): 412-422; B. P. Marx.S. Doron-Lamarca, S. P. Proctor, and J. J. Vasterling, “The Influence of Pre-Deployment Neurocognitive Functioning on Post-Deployment PTSD Symptom Outcomes among Iraq-Deployed Army Soldiers,” Journal of the International Neuropsychological Society 15, no. 6 (2009): 840852; Vasterling et al., “Neuropsychological Outcomes”, S. Maguen et al., “Description of Risk and Resilience Factors among Military Medical Personnel before Deployment to Iraq,” Military Medicine 173, no. 1 (2008):1-9; E. A. Stanley et al., “Mindfulness-Based Mind Fitness Training: A Case Study of a High-Stress Predeployment Military Cohort,” Cognitive and Behavioral Practice 18, no. 4 (2011): 566-576; A. P. Jha et al., “Examining the Protective Effects of Mindfulness Training on Working Memory Capacity and Affective Experience,” Emotion 10, no. 1 (2010): 54-64; A. P. Jha et al., “Minds ‘at Attention’: Mindfulness Training Curbs Attentional Lapses in Military Cohorts,” PLOS One 10, no. 2 (2015): e0116889; A. P. Jha, A. B.Morrison, S. C. Parker, and E. A. Stanley, “Practice Is Protective: Mindfulness Training Promotes Cognitive Resilience in High-Stress Cohorts,” Mindfulness 8, no. 1 (2017): 46-58.
[19]Scaer, The Trauma Spectrum, 62-64; Sapolskv, Why Zebras Don't Get Ulcers,chap. 1; Kavanagh, Stress and Performance, 31.
[20]S. J. Lupien et al., “Cortisol Levels during Human Aging Predict Hippocampal Atrophy and Memory Deficits,” Nature Neuroscience 1 (1998): 69-73;Sapolsky, Why Zebras Don't Get Ulcers, chap. 10. 幸运的是,其他有关动物和人类的研究表明,皮质醇升高的影响不会造成永久性伤害。当皮质醇水平回到正常状态时,海马体也会开始恢复正常大小,参见:McEwen and Lasley, The End of Stress as We Know It, chap. 7. 升高的皮质醇水平与创伤后应激障碍之间的关系并没有那么明确,很多患有创伤后应激障碍的人的皮质醇水平有时候较低,但他们的HPA轴对应激反应依然高度敏感,因此研究人员通常认为创伤后应激障碍导致的海马体体积损失是不可逆的。然而一些研究表明,创伤后应激障碍导致的海马体体积损失是可以得到还原的,参见:E. Vermetten et al., “LongTerm Treatment with Paroxetine Increases Verbal Declarative Memory and Hippocampal volume in Posttraumatic Stress Disorder,” Biological Psychiatry 54 (2003): 693-702; Bremner, Does Stress Damage the Brain?, 60-62, 115119. 其他研究则指出了相反的因果关系:在经历压力性事件之前,海马体和记忆方面出现的小问题会增加遭遇创伤后出现应激障碍的风险,例如,在经历童年期不良经历之后,参见:Marx et al., “The Infuence of Pre-Deployment Neurocognitive Functioning”; R. A. Parslow and A. F. Jorm “Pretrauma and Posttrauma Neurocognitive Functioning and PTSD Symptoms in a Community Sample of Young Adults,” American Journal of Psychiatry 164, no. 3 (2007): 509-515; Sapolsky, Why Zebras Don't Get Ulcers, chap. 10, esp. 222。
[21]K. Cho, “Chronic ‘Jet Lag’ Produces Temporal Lobe Atrophy and Spatial Cognitive Deficits,” Nature Neuroscience 4 (2001): 567-568.
[22]E. S. Brown and P. A. Chandler, “Mood and Cognitive Changes during Systemic Corticosteroid Therapy,” Primary Care Companion for Journal of Clinical Psychiatry 3, no. 1 (2001): 17-21; S. J. Lupien and B. S. McEwen,“The Acute Effects of Corticosteroids on Cognition: Integration of Animal and Human Model Studies,” Brain Research Reviews 24, no. 1 (1997): 1-27.
[23]Sapolsky, Why Zebras Don't Get Ulcers, 221.
[24]Bremner, Does Stress Damage the Brain? 60-62, 115-119.
[25]Bremner, Does Stress Damage the Bnain?, chap. 4; Kavanagh, Stress and Performama, 16-19; McEwen and Lasley, The End of Sress as We Know It,chap. 7.
[26]Daniel J. Siegel, The Developing Mind: How Relationships and the Brain Interact to Shape Who We Are (New York: Guilford, 1999), 253; Ogden et al.,Trauma and the Body, chap. 2.
[27]Porges, The Polyvagal Theory, chap. 1; Ogden et al., Trauma and the Body,chap. 2; Bremner, Does Sress Damage the Brain?, chap. 4; McEwen and Lasley, The End of Stress as We Know lt, chap. 7; Sapolsky, Why Zebras Don't Get Ulcers, chaps. 10, 15.
[28]Kavanagh, Stress and Performance,16-17.
[29]Porges, The Polyvagal Theory, chap. 1; E. A. Scanley, “War Duration and the Micro-Dynamics of Decision-Making under Stress,” Polity 50, no.2 (2018):178-200; J. Renshon, J. J. Lee, and D. Tingley, “Emotions and the MicroFoundations of Commitment Problems,” International Organization 71, no.S1 (2017): S189-S218.
[30]Kavanagh, Stress and Performance, 17-18; Karl E. Weick, Sensemaking in Organizations (New York: Sage, 1995), 129.
[31]Kahneman, Thinking, Fast and Slow; Elizabeth A. Stanley, Paths to Peace: Domestic Coalition Shifts, War Termination and the Korean War (Stanford,CA: Stanford University Press, 2009), chap. 2; Kavanagh, Stress and Perfornamce, 17-19; Scott Sigmund Gartner, Strategic Assessment in War(New Haven, CT: Yale University Press, 1999).
[32]R. F. Baumeister et al., “Bad Is Stronger Than Good,” Review of General Psychology 5, no. 4 (2001): 323-370; P. Rozin and E. B. Royzman,“Negativity Bias, Negativity Dominance, and Contagion,” Personality and Social Psychology Review 5, no. 4 (2001): 296-320.
[33]Stanley, “War Duration”; Kavanagh, Stress and Performance, 17-20;Kahneman, Thinking, Fast and Slow.
[34]Scaer, The Trauma Spectrum, 58-59, 132-133.
[35]Scaer, The Trauma Spectrum, chap. 3; Ogdenet al., Trauma and the Body, 2022, 86-87.
[36]Scaer, The Trauma Spectrum, 42, 95, chap. 3; Ogden et al., Trauma and the Body, 34-36, 86-87.
[37]Scaer, The Trauma Spectrum, chap. 3, 42; Ogden et al., Trauma and the Body, 18-23, 86-87, 104-105; B. A. van der Kolk, “Clinical Implications of Neuroscience Research in PTSD,” Annals of the New York Academy of Sciences 1071, no. 1 (2006): 277-293.
[38]Scaer, The Trauma Spectrum, 59-64; Robert C. Scaer, The Body Bears the Burden: Trauma, Dissociation, and Disease, 3rd ed. (New York: Routledge,2014), 91-95.
[39]Bessel A. van der Kolk, The Body Keeps tbe Score: Brain, Mind, and Body in the Healing of Trauma (New York: Penguin, 2015), chap. 11; Sapolsky, Why Zebras Don't Get Ulcers, 320-323; Scaer, The Body Bears the Burden, 91-95.
[40]Scaer, The Trauma Spectrum, 62-67.
[41]Scaer, The Trauma Spectrum, 62-64.
[42]Ogden et al, Trauma and the Body, 18-23.
[43]Scaer, The Trauma Spectrum, 62-64; Pat Ogden and Janina Fisher,Sensorimotor Psychotherapy: Interventios Trauma and Attachment (New York: Norton, 2015), 181.
[44]Ogden et al., Trauma and the Body, 7.
[45]McEwen and Lasley, The End of Stress as We Know It, 37-38; Joseph LeDoux, The Emotional Brain: The Mysterious Underpinnings of Emotional Life (New York: Touchstone, 1998); M. R. Delgado, A. Olsson, and E.A. Phelps, “Extending Animal Models of Fear Conditioning to Humans,”Biological Psychiatry 73 (2006): 39-48; A. Feder, E. J. Nestler, and D. S.Charney, “Psychobiology and Molecular Genetics of Resilience,” Nature Reviews Neuroscience 10 (2009): 446-457; M. R. Delgado et al., “Neural Circuitry Underlying the Regulation of Conditioned Fear and Its Relation to Extinction,” Neuron 59 (2008): 829-838; M. R. Milad et al, “Thickness of Ventromedial Prefrontal Cortex in Humans Is Correlated with Extinction Memory,” Proceedings of the National Academy of Sciences 102, no. 30(2005): 10706-10711; D. Schiller et al., “From Fear to Safety and Back:Reversal of Fear in the Human Brain.” Journal of Neuroscience 28 (2008):11517-11525.
[46]Ogden et al., Trauma and the Body, 10-11.
[47]van der Kolk, The Body Keeps the Score, 182; van der Kolk, “Clinical Implications of Neuroscience Research in PTSD,” 281-282; Ogden et al.,Trauma and the Body, 23-24, 37.
[48]Ogden et al., Trauma and the Body, 24.








本书评论