
体内糖的调节
碳水化合物是人体内的一种重要的燃料来源,但其数量有限。休息状态下,肝脏、胰腺和其他器官一起将血糖水平控制在一个较小的范围内,以满足不同身体组织对碳水化合物的需求。因为骨骼肌中存储的有限的糖原是肌肉收缩的重要能量来源,所以休息状态下人体很少用到这种碳水化合物。进食后,人体尽可能多地以糖原的形式存储碳水化合物,同时刺激碳水化合物燃料的使用,使血糖水平回归正常。在禁食状态,人体将调动葡萄糖前体物质来实现肝脏糖原异生(肝脏糖异生),同时促进脂肪氧化以便获取能量来节省碳水化合物燃料。
在运动和比赛期间,人体通过增加碳水化合物和脂肪的分解,以及提高肝脏糖异生的速率来维持血糖水平。运动中碳水化合物和脂肪的动员程度取决于多个因素,但是最为重要的因素是运动本身(例如参与运动的肌肉总量和肌肉收缩的强度)。
血糖稳定
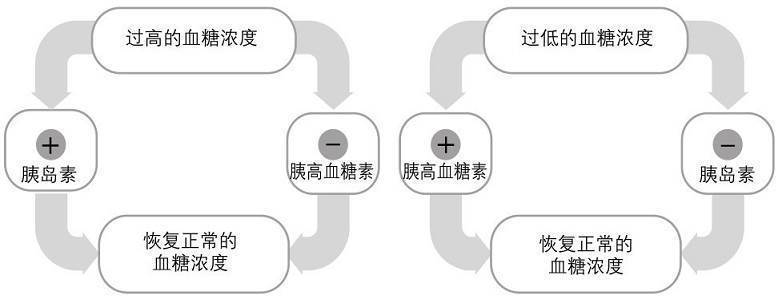
一般情况下,一个成人身体里的总血量大约为5升。在这5升总血量中,大约有5克葡萄糖。从食物中摄取碳水化合物、肝糖原分解(肝糖分解)和糖异生都有助于保持血糖水平。在禁食阶段,糖异生对血糖水平的保持作用更大。休息状态下,肌肉内的葡萄糖和糖原使用率很低。血浆胰高血糖素和胰岛素的平衡对血糖和身体组织的糖原利用具有最强的调节功效。当血糖低于正常水平时,胰腺的α细胞就会分泌胰高血糖素。胰高血糖素是一种碳水化合物调动激素,其可以促进肝脏的糖异生和肝糖分解,从而使血糖水平回归正常(见图2.2)。当血糖水平在进食后超过正常值,胰腺的β细胞就会分泌胰岛素。胰岛素降低血液中葡萄糖的方式有两种,包括增加胰岛素敏感的组织(主要是骨骼肌和脂肪组织)的血流量和刺激糖分子扩散到对胰岛素敏感的细胞中来。胰岛素也可以刺激细胞的能量代谢,氧化碳水化合物,加快以糖原的方式存储葡萄糖,并抑制肝脏和骨骼肌的糖原分解和肝脏糖异生。在训练实践的角度,确保这些系统的正常工作对于保持血糖水平非常重要,因为有氧耐力会随血糖水平的下降而下降。
糖原合成
糖原存储在骨骼肌和肝脏中。肌糖原是高强度有氧或无氧运动的一种非常重要的能源物质。肝糖原降解成葡萄糖,然后被输送到血液中,以便在有氧耐力运动中维持血糖水平。本节将解释糖原是如何被合成的。
在糖原合成过程中,细胞内的葡萄糖要几经转变才能生成尿苷二磷酸(UDP)-葡萄糖(Leloir,1971)。这一反应需要经历三个步骤:
1.葡萄糖在进入细胞的过程中被己糖激酶催化成葡萄糖-6-磷酸。
2.葡萄糖-6-磷酸通过葡萄糖磷酸变位酶转化为葡萄糖-1-磷酸。
3.在由尿苷二磷酸-葡萄糖焦磷酸化酶催化的反应过程中,葡萄糖-1-磷酸和尿苷三磷酸合成尿苷二磷酸-葡萄糖。
形成的尿苷二磷酸-葡萄糖可以促进糖原分子的增长。这一过程受到糖原合成酶的催化作用。这时,只要多糖链含有四个以上的葡萄糖残基,就会使其再增加一个葡萄糖单位。糖原不单单是一长串重复的葡萄糖合成物,它还是一种多分支的聚合物。这种分支很重要,因为它能增加糖原的溶解度,也能够促进糖原的快速合成与降解(在高强度运动中,这有助于提供更多葡萄糖,使其进行糖酵解来产生能量)。
➤ 磷酸化——将磷酸基团加在另一个分子上的过程。磷酸化可以激活和去激活很多蛋白酶。
糖原分解
一旦运动过程中糖原发生降解,这就说明人体需要ATP来为骨骼肌收缩提供燃料。糖原分解的目的是释放葡萄糖(特别是葡萄糖-1-磷酸)化合物,并使葡萄糖可以进入糖酵解,加快ATP的生成。
➤ ATP——一种由细胞合成和使用的高能磷酸化合物,用于释放能量供细胞工作所用。
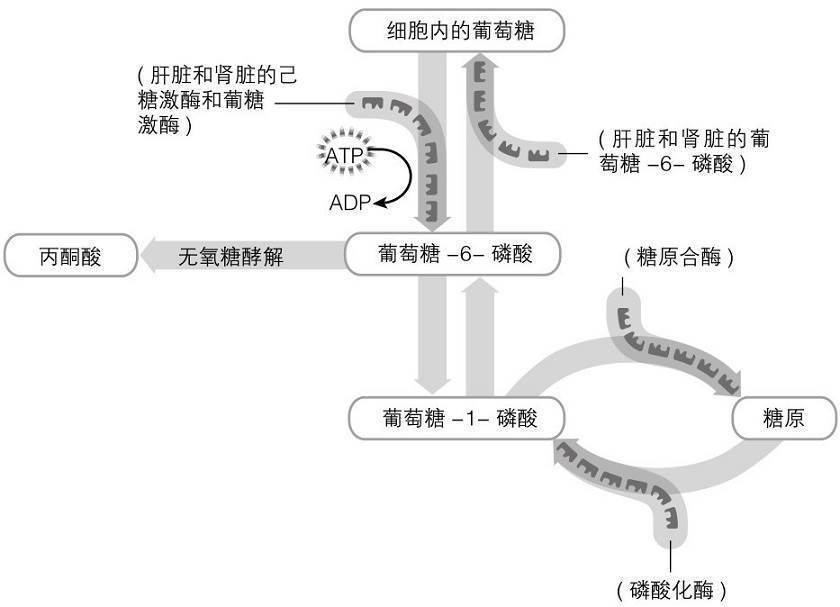
在糖原分解的复杂过程中,糖原分解产生的单个葡萄糖化合物形成葡萄糖-1-磷酸(由磷酸化酶催化而成)。磷酸化酶将糖基残留物从糖原分子的非还原端逐一移除。糖原分解过程中形成的葡萄糖-1-磷酸被葡萄糖磷酸变位酶转化为葡萄糖-6-磷酸。在骨骼肌中,从糖原分解出来的葡萄糖-6-磷酸,与由血液进入细胞的葡萄糖转化而成的葡萄糖-6-磷酸一起,进入糖酵解代谢过程。肝脏,和一部分的肾脏,要么通过糖酵解处理糖原分解的葡萄糖-6-磷酸,要么将糖原分解的葡萄糖-6-磷酸去磷酸化,然后将这些葡萄糖释放到血液中。在细胞葡萄糖代谢过程中(例如糖原合成和糖原分解),葡萄糖-6-磷酸这种媒介在葡萄糖存储和葡萄糖氧化的各种转化过程中发挥中心作用(见图
2.3)。
糖酵解
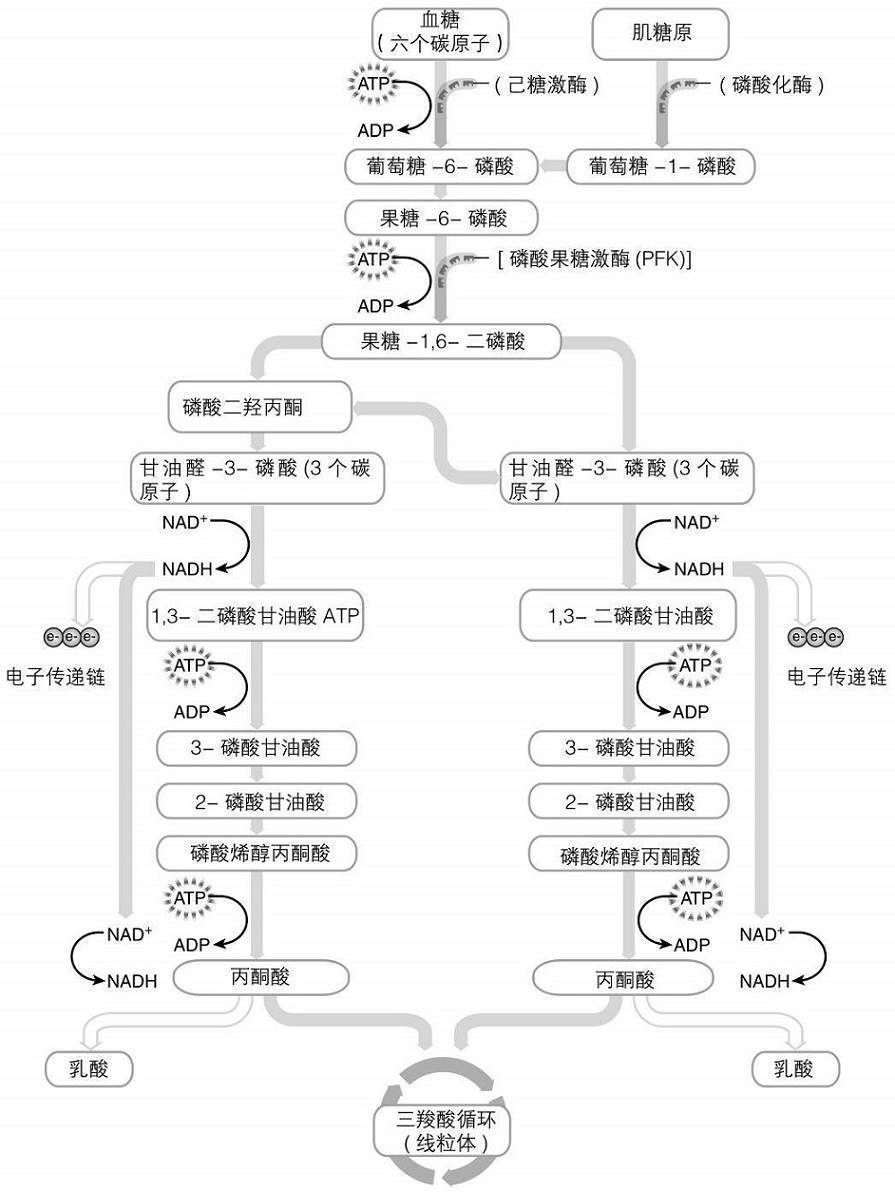
在运动、高强度训练和比赛中,人体需要快速获得ATP以便产生能量。产生ATP最快的方式之一就是糖酵解。简而言之,糖酵解就是分解碳水化合物(例如葡萄糖)来产生ATP的过程。糖酵解发生在肌肉组织的细胞质中。糖酵解的一个重要生理成果就是能相对较快地生成ATP,从而用于肌肉收缩。从图2.4可以看出,糖酵解是一组10个由酶控制的化学反应链。它的起点是一个含有六个碳原子的葡萄糖,终点是两个含有三个碳原子的丙酮酸分子。
糖酵解最后产生的丙酮酸可能有两种命运:转变为乳酸,或进入线粒体。接下来的一节将描述乳酸的产生。进入线粒体的丙酮酸先转化为乙酰辅酶A,然后再进入众所周知的三羧酸循环。三羧酸循环在一系列酶催化的化学反应过程中,进一步使丙酮酸-乙酰辅酶A进行代谢。最终,三羧酸循环里的这些化学反应会生成烟酰胺腺嘌呤二核苷酸(NADH)和黄素腺嘌呤二核苷酸(FADH2)。他们会将电子传递给线粒体中的电子传递链。这些电子传递链促进生成更多的ATP来为骨骼肌收缩提供燃料。但是与糖酵解生成的ATP相比,这种ATP生成的速率较慢。了解糖酵解产生ATP的速率更快这一点是非常重要的,特别是在高强度训练或运动中。这种ATP生成方式主要是由葡萄糖分解来促成的,所以确保膳食中含有充足碳水化合物的重要性也就很容易理解了,这样才能在训练和比赛中为高强度运动提供燃料。
➤ 线粒体——细胞中负责利用氧产生ATP的结构,三羧酸循环、电子传递链和脂肪酸循环均位于线粒体。
乳酸的产生和清除
前文已经提到,糖酵解的终产物是丙酮酸。丙酮酸可以转化为乙酰辅酶A,然后进入三羧酸循环。丙酮酸也可以转化为乳酸。丙酮酸转化为乳酸的这一过程被称为无氧糖酵解。一旦在细胞内生成,乳酸就通过释放氢离子进行快速解离,并降低细胞质的pH值。解离出氢离子后,乳酸变为乳酸根。细胞内的pH值随乳酸的产生而下降,这会对众多代谢环节和肌肉收缩过程产生不利的影响。因此,必须立即减少细胞中的乳酸或者将其排除到细胞外清除。人体在休息和低运动强度时会产生少量的乳酸。大部分乳酸在细胞内就可以很容易地被清除。有些乳酸需要被运输到细胞外进行快速无害的处理。血浆血红蛋白是最重要的细胞外缓冲物。此外,血浆中的碳酸氢根也具有细胞外化学缓冲的作用。在持续高强度的肌肉收缩过程中,肌肉会有疼痛或灼烧的感觉。这主要是由于pH值下降刺激了肌细胞外游离神经细胞末梢。剩下的含有3个碳原子的乳酸根是可以被用作非运动肌、心肌,甚至是参与运动的肌肉本身的一种潜在燃料(Van Hall,2000)。在无氧阈所对应的中高强度运动中,乳酸的生成速率超过了细胞内的缓冲能力,这造成多余的乳酸就被转移到细胞外。随着运动强度的增加,血液中的乳酸水平会快速增加。高强度运动中产生的过多乳酸对肌肉做功能力有负面作用。但是这种代谢副产品的生成有助于促进短时间内碳水化合物经无氧代谢生成ATP。
➤ 肌浆——肌纤维的细胞质。
➤ 无氧阈——该术语通常用来指运动时血乳酸浓度快速增加的转折点所对应的摄氧量水平。
疲劳,被定义为不能维持预定的功率输出或运动强度。在短时高强度无氧运动中,疲劳产生的一部分原因是参与运动的肌肉中乳酸的堆积(导致pH值下降)。持续的高强度运动会快速耗尽参与运动的肌肉中的糖原。细胞间糖代谢底物的减少和血糖获取速率的受限使得肌肉迅速疲劳,同时无氧系统维持快速ATP再合成的能力也在下降。随着有氧供能对整体运动所需ATP的贡献越来越大,肌肉收缩可以以较低强度继续进行。
➤ 底物——酶可以作用的分子。
尽管乳酸的堆积与疲劳有关,但没有任何饮食方法能降低运动中乳酸的产生。相反,适当的训练能使运动员即使在高强度运动时乳酸也可以处于相对较低的水平。一般来说,运动员的饮食中包含最佳剂量的碳水化合物是很重要的,因为这使运动员能够完成高强度的训练。这种训练将导致机体产生代谢适应,改变能量物质的利用(如脂肪)来满足运动时的能量需求。
身体中的糖供能受其储备量的影响。血糖浓度的变化为肝糖原的释放提供反馈调节,血糖浓度上升会抑制运动中肝糖的释放。碳水化合物的储备情况还会通过降低细胞中脂肪酸的动员和氧化来限制脂肪代谢(Spriet,1998)。简而言之,当ATP需求大时,脂肪酸氧化因供能速率太慢而无法满足能量所需,这样会导致线粒体NADH和乙酰辅酶A负荷的加大,必要时会迫使糖通过产生乳酸的方式来维持无氧糖酵解。








本书评论